Burrack, North Carolina State University, Bugwood.org
The spotted wing drosophila (SWD), Drosophila suzukii, is an invasive vinegar fly. The fly is small (about 2-3mm in size) with a pale body and bright red eyes. Male SWD have a black spot near their wingtip and black bands on their front legs, which may help to distinguish them from other look-alike vinegar flies. This species of fly was first observed in the United States in California in 2008; it was found in New England in 2011. SWD lay their eggs in ripe or ripening fruit and berries. When the larvae hatch, they consume the fruit leaving it deformed. Hannah
Habitat
SWD affects many fruit and berries, including cherries, peaches, plums, pears, apples, raspberries, blackberries, strawberries, grapes, blueberries, and tomatoes. The insect also can infest non-commercial hosts such as crabapple, autumn olive, Japanese yew, beach plum, and wild rose. It not only infects already ripened fruit, but can affect fruit in the earlier stages of development as well. SWD are seen more in moderate climates and prefer the early summer or fall when temperatures are cooler (approximately 68° F). Adults will hibernate when temperatures drop to 40°F. In freezing temperatures, the larvae usually cannot survive.
Origin
Spotted wing drosophila are native to Southeast Asia.
Introduction and Spread
Various fruit crops will develop and ripen at different times throughout the growing season. The SWD will take advantage of this and move from one crop to another as the season progresses. Additionally, infested fruit may be shipped from different parts of the country and sold at markets, which increases the spread of SWD. This insect has a high reproductive rate. Within one year, the SWD can complete at least 15 generations. During the 2-9 weeks time frame that adults live, a female can lay 100-600 eggs in fruit.
Impact
When female SWD lay their eggs on the fruit, a small, barely visible scar is left on the fruit’s surface. Within a couple hours or a few days the larvae may begin to hatch and feed on the fruit from the inside, which eventually causes the fruit to collapse and look deformed. Fruit may start looking misshapen within 2-13 days after becoming infested. Secondary pests, such as molds and bacteria, may also further infect the fruit, exacerbating the damage. Since SWD look very similar to other common vinegar flies, many populations of SWD go unnoticed and untreated until a large infestation occurs.
In 2009, yield loss from SWD was estimated to be in a range from zero up to 80% (Walsh et al. 2011). SWD is a big concern in areas located on the western coast of the United States, such as California, Oregon, and Washington. There, a large proportion of the nation’s commercial production of raspberry, blackberry, cherry, strawberry, and blueberry occurs. In the Northeast, SWD has affected raspberry and blackberry the most. Peaches and grapes are also affected in the Northeast. It is still unknown how this species will overwinter and if earlier season crops are of greater risk to be infested in the future (Demchak et al. 2012).
D. suzukii damage to cherry fruit. Photo: G. Arakelian, cisr.ucr.edu.D. suzukii larvae inside a cherry. Photo: UC Statewide IPM Program, University of California, cisr.ucr.edu.
Identification
SWD is about 2-3 mm in size, with bright red eyes. The majority of the insect’s body is a pale brown with black horizontal stripes on the abdomen region. You can distinguish a male SWD from other vinegar flies in the Drosophila genus by a dark spot along the front edge of the wing near the wingtip. This spot may be missing on some SWD males, but another defining characteristic for males is the presence of two black bands on each front leg. Female SWD tend to have a saw-like ovipositor, larger than that of other species. Otherwise they look very similar to common vinegar flies. The white larvae of SWD are small and cylinder-shaped.
Male Drosophilia suzukii. Photo: G. Arakelian, cisr.ucr.edu
Prevention
Growers of fruits that are commonly affected by SWD are encouraged to inspect their crops frequently. Screens or growing inside a greenhouse have been used for individual plant protection to exclude the flies in some cases. However, this method may not be as suitable or practical for many growers and producers. Bait traps or lures to detect fruit flies may also be utilized to monitor the presence of SWD. Since females have fewer distinguishing characteristics than males, it is recommended to monitor the male populations to avoid confusion with other species. Traps should be placed in a field right before the fruit starts to turn color. For more information on making your own traps and placement of traps, refer to the Pennsylvania State University monitoring fact sheet, (Demchak et al. 2012).
Control
To control already-established infestations of SWD, remove all ripe and cull fruit when harvesting. Also, try to pick the fruit as soon as possible and do not leave harvested fruit exposed for long periods of time. Any fruit that has become infested should be promptly removed and disposed of properly. All damaged fruit should be buried, covered with plastic, or disposed of in a closed container. Leaving the infested fruit to decompose in the field where they dropped will give eggs and larvae still present on the fruit a chance to fully develop into mature insects causing the population on SWD to increase.
There are some biological control agents, such as parasitoids that affect species within the Drosophila genus (Dubuffett et al. 2009). Researchers at Oregon State University are exploring biological control methods specifically for SWD. However, it is still unknown if these predators are a reliable biocontrol method at this time. Some pesticides that are approved may be applied across fields for SWD control. These treatments may need to be reapplied almost every 1-2 weeks. Three groups of pesticides, the pyrethroids, spinosyns, and organophosphates have been found to be effective. Since SWD has such a high reproductive rate, with multiple generations per year, it is suggested to use a variety of pesticides to reduce chemical resistance evolving within this species (Demchak et al. 2012). Contact your local Cornell Cooperative Extension office for more information on pesticides before use and always follow the directions on the label.
Occurrences
SWD has been documented in Japan, China, Korea, Thailand, India, Spain, and Hawaii. SWD was first observed in California in 2008. In 2011, SWD was found in the Northeast in Pennsylvania.
New York Distribution Map
This map shows confirmed observations (green points) submitted to the NYS Invasive Species Database. Absence of data does not necessarily mean absence of the species at that site, but that it has not been reported there. For more information, please visit iMapInvasives.
Asiatic clam, Prosperity clam, Pygmy Clam, Golden clam, Good luck clam
Origin and Spread
The Asian Clam Corbicula fluminea (Müller) is native to the fresh waters of eastern and southern Asia. It was likely introduced to the West Coast of North America around 1930, initially assumed to have been imported as a food source for the immigrating Chinese population (USACE ERDC 2007). Alternatively, it may have come in with the importation of the Giant Pacific oyster also from the Asia (Foster 2012). Live Asian clams were first detected in US waters in 1938 in the Columbia River, Washington; the species quickly spread across the continent and is currently found in 44 states. Corbicula was detected in the Ohio River in 1957 and continues to spread through drainages in the Midwest and Northeast. In New York State, the Asian clam has been collected in the running waters of central and western portions of the state, the Erie Canal from Lockport east to Clyde (shells only, colonization status unknown), Canandaigua Lake Outlet (shells only, colonization status unknown), Canandaigua, Keuka, Otisco, Owasco, and Seneca Lakes in the Finger Lakes, the Hudson River from Troy to Newburgh, the Wallkill River (colonization status unknown), the Champlain Canal near Fort Edwards, Lake George, and in Massapequa Lake and a number of other ponds and streams and the Massapequa Reservoir on Long Island.
The exact mechanism for secondary dispersal of Corbicula throughout North America is unknown, but likely involves human activity, including bait bucket introductions, accidental introductions associated with imported aquaculture species, and intentional introductions by people who buy them as a food item in markets (Foster 2012). Larval clams can attach to vegetation, floating debris for long distance dispersal. There is no peer-reviewed data demonstrating long distance overland travel by juveniles as a result of byssal attachment to trailered boats (McMahon 2012). Juvenile Corbicula are more likely to be carried in bilge and livewell water in boats and on vegetation attached to anchors and trailers or in sediments left on anchors (McMahon 2012).The only other significant dispersal agent is thought to be passive movement via water currents. There remains some question regarding transport by waterfowl. Based upon Foster (2012), birds are not considered to be significant distribution vectors. Sousa, et. al. (2008) cites a number of credible peer-reviewed sources indicating that transport on the feathers and feet of water birds is a secondary transport vector.
Identification
The species has a typical oval-triangular clam shape, with a dorsal “beak” or umbo at the peak of the shell. The outside of the shell (periostracum) is olive, or yellowish to black-brown in color, with 1-3 brown/purple colored radial bands (particularly in juveniles) and white erosion rings near the umbo. As the clam ages, the periostracum becomes darker in color. There are also distinctive, thick, concentric growth rings on the periostracum. The inside layer of the shell, the nacre (or “mother of pearl layer) is typically white-bluish white in color. Inside each shell half (or valve), there are also 1-2 pair of small, elongated and , finely serrated lateral “teeth” that extend on either side from the umbo part way down on the inside edge of each valve. Also, the interior of each valve, immediately under the umbo, there are 3 cardinal “teeth” (MacNeill 2012). The clam most closely resembles native sphaerid (fingernail) clams, however, sphaerid clams are smaller (6-14 mm), more oval in shape, cream colored, have fine growth rings, lack serrations on the lateral teeth and are found completely buried in the sediment. These relatives of Corbicula are also found in slow flowing waters with poorer water quality.
Corbicula fluminea External (left valve) and Internal (right valve) body features
Corbicula fluminea identication key
Biology
Corbicula burrows into the bottom sediments of streams and lakes and has the ability to feed from both the water column and the substrate. It uses its siphon to filter feed suspended particles (particularly phytoplankton) from the water and its fleshy foot appendage to pedal feed on detritus in the sediment. Corbicula can live in a variety of substrates, but prefers sand and gravel, over silt hard surfaces (McMahon 1999).
Corbicula is less tolerant than native mussels to environmental fluctuations. It is extremely sensitive to low oxygen conditions, and consequently its distribution is restricted to well‐oxygenated streams and lake shallows. In its native semi‐tropical/tropical habitat, the Asian clam is rarely exposed to temperature extremes. Its northern distribution in North America is thought to be limited by a 2°C lower lethal limit, and reproduction requires sustained water temperatures of 15°‐16°C. Asian clams have been known to find temperature refuges in cooler waters heated by power plant discharge. Upper lethal limit is believed to be 30°-35°C (Foster 2012, MacNeill 2012). Corbicula has been found to be resistant to desiccation and can survive periods of low water in damp sand or mud (USACE ERDC 2007).
Adult Corbicula are simultaneous hermaphrodites (both male and female) that are capable of both cross and self‐fertilization; thus, it takes only 1 individual to start a population. [Note: some literature indicates that Corbicula also exists as male and female sexes; this idea might be a result of observation of cross fertilization between hermaphrodites.] Adults can live 3‐4 years, and typically reproduce two times a year, although some populations have been observed reproducing more often under optimal situations. A single adult can produce 1000 – 100,000 juveniles per year. Egg fertilization is internal and the larval clams are brooded on the gill where they transform into juveniles in about 4-5 days (MacNeill 2012). Juveniles are tiny (0.25mm) and are capable of long distance dispersal via stream transport and water currents (using a mucous “balloon” or “parachute” {MacNeill 2012}), or hitchhiking on animals, floating objects, or vegetation (to which they can attach via a byssal thread). The juveniles transform into a pediveliger (shelled juvenile equipped with a foot) and settle to the bottom at about 0.25 mm in size. Pediveligers can crawl around along the bottom and seek firm substrates where they attach (at about 1.0 to 1.5 mm in size) using a temporary byssus which eventually disappears (MacNeill 2012). Juvenile clams can reach maturity in 3‐6 months or about 6-10 mm in size, and reach 10 to 30 mm in size during their first year depending on food availability and temperatures (MacNeill 2012).
Corbicula can rapidly grow into dense populations (> 2,000 per square meter), but are prone to rapid die‐offs with sudden changes in temperature (hot or cold) and low oxygen. However, their life history traits (i.e., quick maturity, high fecundity) enable rapid re‐colonization and population recovery, even after near extirpation. Additionally, these traits allow the Asian clam to successfully colonize habitats disturbed by human activity (e.g., channels and impoundments) that are unsuitable for native mussels.
Ecology
Like other bivalves, Corbicula is a filter-feeder on microscopic plants, animals (including bacteria) and in the water column or in the sediments. Among other freshwater bivalves it has the highest rates of filtration rates (up to 1.3 liters/hr/clam), food consumption and growth of any species (MacNeill 2012). It is also extremely efficient in channeling consumed food for growth and reproduction. Its ability to reproduce rapidly, coupled with low tolerance of cold temperatures, can produce wide swings in population sizes from year to year in northern water bodies. Both yellow and brown morphs are simultaneous hermaphrodites and brood their larvae in the inner demibranchs (Foster 2012). The life span is about one to seven years. No large-scale geographic features function as dispersal barriers.
Impacts
Ecological Impacts
Like other invasive mussels (e.g. zebra and quagga mussels), Corbicula is highly successful coupling the nutrient and energy flows that occur in the water column and bottom sediments. With a high filtering capacity and population density, Corbicula filters out phytoplankton and other particles suspended in the water that are also important food sources for other filter‐feeding organisms. Unlike zebra and quagga mussels, Corbicula also uses its pedal foot to feed on organic material and tiny organisms (microbes, protists, meiofauna) in the sediment (Hakenkamp et al. 2001). Whether Corbicula depletes these food resources to the extent that it negatively affects other organisms (particularly native unionid and sphaeriid mussels) remains an open question (Strayer 1999).
Corbicula can affect aquatic ecosystem processes in other ways. Bivalves, particularly when in dense populations, excrete significant amounts of inorganic nutrients, particularly nitrogen that, in turn, can stimulate the growth of algae and macrophytes (Lauritsen and Mozley 1989, Sousa et al. 2008). Additionally, Asian clam mass mortality events that occur in the summer followed by the release of nutrients via decomposition may also have negative effects on water quality. The shells of dead Asian clams can also provide a hard substrate on soft sediments, creating new habitat for other species that prefer hard substrates (e.g., zebra mussels).
It should be noted that to date, there are few studies on the ecological impacts of the Asian clam on native biota (McMahon 1999). In fact, most studies examine a water body after Corbicula invasion, with no comparable information on the biota or environmental conditions pre‐invasion.
Moreover, recent justifications for expensive and intensive management actions (e.g., Lake Tahoe Asian Clam Response) overstate evidence in the scientific literature. For example, in “Asian clam (Corbicula fluminea) of Lake Tahoe: Preliminary scientific findings in support of a management plan,” Wittman et al. (2008) state that:
“Asian clam is known to aggressively outcompete native invertebrate communities.”
Here the report authors incorrectly interpret and cite the findings of Karatayev et al. (2003). Karatayev and colleagues studied Corbicula in a Texas reservoir. Although the Asian clam dominated the total animal biomass of the reservoir sediments (up to 95%), it was not associated with declines in native biodiversity. In fact, it was found to co‐occur with an abundant population of native unionid mussels.
Studies claiming that the Asian clam has an impact on native bivalves (particularly unionids) are often anecdotal and only report the spatial distribution of bivalves after invasion (Strayer 1999). They assume that non‐overlapping distributions of Corbicula and native mussels indicate that Corbicula have outcompeted the native species. As Strayer (1999) points out, this is just one possible explanation. Corbicula could also prefer different habitat than native mussels (e.g. sandy vs. silt/gravel). Native mussels have long experienced declines due to human‐induced changes to habitat (pollution, land use change, channelization), and it is difficult to tease apart these changes versus direct impacts of invading Corbicula that also happen to do well in disturbed habitats. Thus, while it is quite possible that in some cases Corbicula has a direct, negative impact on native biota, more studies that monitor changes in mussel populations over time and that directly evaluate competitive interactions are warranted.
Economic Impacts
Asian clams can colonize the intake pipes of water treatment systems and power stations. Unlike zebra mussels, Corbicula do not attach to the hard substrate. Rather, juvenile clams pass through filter screens/strainers and settle on the floors of intake pipes where low flow allows silt and sand to settle. The clams reproduce in situ and continue to accumulate in pipes and are transported deeper into the system. Corbicula fouls the pipes, blocking structures with shells, altering flow, and increasing sedimentation rates (McMahon 1999).
Asian clam shells that accumulate in beach or swimming areas will also impede recreation and tourism.
Control and Management
Little to no research regarding control practices have been published in the peer‐reviewed scientific literature. The Lake Tahoe team has submitted a manuscript for publication to the journal Biological Invasions on the use of bottom barriers to control Corbicula and it remains in the peer review process. Additionally, the New York Invasive Species Research Institute had difficulty locating research-based Best Management Practices for Corbicula.
To date, given the life history traits of Corbicula that make it a successful invader and the widespread distribution of the species in North America, eradication is typically not a viable option. As with many aquatic invaders, emphasis ought to shift from eradication to containment and spread prevention.
New York Distribution Map
This map shows confirmed observations (green points) submitted to the NYS Invasive Species Database. Absence of data does not necessarily mean absence of the species at that site, but that it has not been reported there. For more information, please visit iMapInvasives.
Acknowledgment
New York Invasive Species Information wishes to acknowledge the work of Dr. Holly Menninger, former director of the New York Invasive Species Research Institute. Dr. Menninger’s literature review formed the basis for this species profile and provided much of the text published above.
European crane flies (Tipula paludosa), also known as leather jackets during the larval stage, are an invasive insect that have become established in the northwestern United States, eastern Canada, and New York. Crane flies look like large mosquitoes, but exploit a different ecological niche and do not bite humans. Crane flies are most problematic to the turf grass industry, especially on golf courses. This is because the flies lay their eggs in the soil for larvae to feed. Tipula paludosa as well as Tipula oleracea are exotic European crane flies that are present in New York.
Adult European crane fly. Photo: Canadian Forest Service Archive, Canadian Forest Service, Bugwood.org
Origin
European crane flies (T. paludosa) are native to northwestern Europe.
Habitat
European crane flies thrive in moist soils and prefer mild winters and cool summers. In Quebec, Canada, T. paludosa had higher larval abundance in silt or clay soils rather than sandy soils (Taschereau et al. 2009). They have been found in many different types of turf grass (at homes and golf courses), sod farms, and grass fields. The flies also favor areas with thatch buildup.
Introduction and Spread
European crane flies were most likely introduced through the transport of infested materials. During the summer months, T. paludosa larvae will reside below the soil surface and pupate. By end of August or early September, the adults emerge, mate, and lay 200-300 eggs within 24 hours. Once the eggs hatch, the larvae will feed throughout the fall and spring. T. paludosa complete one generation per year and T. oleracea will complete two generations per year with adults emerging in the spring in addition to the fall. T. oleracea females are better fliers, which could contribute to their eggs being more dispersed than T. paludosa eggs (Peck 2006).
Impacts
European crane flies have a large impact on the turf grass industry. The larvae may scalp lawns during foliar feeding and damage the root systems as well. Dawson et al. (2004) found that larvae of crane flies significantly impact shoot and root biomass and the microbial composition of the soil. Damaged lawns may appear yellow or have bare spots. Crane fly larvae have the potential to affect cereals and produce crops, nursery stocks, bare root and container stock, and flowers as well. The highest damage intensity from T. paludosa has been observed in late spring. Additionally, some natural predators, such as birds and skunks, may disrupt lawns when foraging for larvae in the grass. Natural predators feeding on larvae and swarms of mature adult crane flies maybe become nuisance species for home owners.
European crany fly adult on ground foliage. Photo: Whitney Cranshaw, Colorado State University, Bugwood.org
Identification
European crane flies are about a half-inch long, with six long legs. A distinguishing characteristic of adult European crane flies is a dark-colored band on the leading edge of the wing next to a light colored band. The rest of the wing has no pigmented areas, which is useful for distinguishing them apart from some native species. Eggs of European crane flies are black and oval shaped and can be found near the soil surface. The larvae are gray-brown and worm-like. Larvae have a tough skin with visible veins. The empty pupal cases may be seen in low turf and look like small grey-black twigs.
Wing of adult T. paludosa. Photo: John Sankey, web.ncf.ca/bf250/gardendip.html
Prevention and Control
Monitoring: Surveying for crane fly larvae should be performed in the early spring by observing turf damage. To survey, take core samples or turn over the top 1-2 inches of sod in one square foot plots and count the larvae. Pouring warm water with dish soap on a mowed plot will also make the larvae emerge if you prefer not to dig the sod up; however, this method may not be as successful. In the fall, surveying for the pupal cases can be performed in low cut grass. In the fall, adults should be noticeable especially in large infestations. Since T. paludosa adults do not fly far, the surrounding areas should be surveyed for eggs and larvae in the current and future years. If you are unsure if you are looking at an invasive European crane fly, collect samples and take them in to your local Cornell University Cooperative Extension office or send them to a specialist or diagnostic lab.
Manual and Mechanical Control: Maintaining proper turf grass health may help to allow the grass to recover from damage faster. Applying fertilizers in the fall will help maintain turf quality. Increasing soil aeration and dethatching in the spring may help reduce crane fly populations. Also, because larvae thrive in moist soils, timing of irrigation may be critical during the oviposition stage of the lifecycle and throughout the fall season. Draining soils during the critical period of the life cycle may be beneficial to reduce fly populations in infested soils. Eggs that are laid at the soil surface and the larvae that emerge can be raked up and destroyed to prevent future damage from occurring.
Biological Controls: Crane flies have some natural predators, such as various species of birds or microorganisms. Beauveria bassiana is a fungus that has been seen to attack crane flies. Nematodes applied in the spring have been effective in some areas as well (mainly the Northwestern United States) in 55 degree temperatures and irrigated soil. For some fly populations, manual and biological control methods are environmentally preferred.
Chemical Controls: Chemical controls for crane flies are most effective if applied in the fall during the egg laying period, but may be used in the spring as well. Imidacloprid, trichlorfon, carbaryl and chlorpyrifos are effective against crane flies. Contact your local extension office for more information on chemical pesticides and always read the instructions on the labels. For more information on potential lawn and turf solutions: http://www.northeastipm.org/schools/outside/low-maintenance-turf/ http://www.organiclandcare.net/store/nofa-organic-lawn-and-turf-handbook
To prevent additional spread of crane flies, movement of sod, container stock, and other turf grass materials should be limited.
New York Occurrences
European crane flies (Tipula Paludosa) were first detected in New York State in 2004. This species has been found in Erie, Monroe and Niagara counties in New York and is most prevalent in the western half of the Erie Canal corridor. A similar species of crane fly, Tipula oleracea, is also present in Monroe, Niagara, Ontario, Onondaga, Oswego, Seneca, Wayne, Nassau and Suffolk counties in New York.
Other Occurrences
T. paludosa has been documented in the Northwestern United States, New York and Eastern Canada.
Pale swallow-wort or dog-strangling vine (Vincetoxicum rossicum; syn. Cynanchum rossicum) and black swallow-wort or black dog-strangling vine (Vincetoxicum nigrum; syn. Cynanchum louiseae) are herbaceous invasive plant species. They were originally placed in the milkweed family, Asclepiadaceae, but in recent years the Asclepiadaceae has been recombined with the periwinkle family, Apocynaceae. Pale and black swallow-wort are two distinct species, but do have a similar growth habit and many similar physical characteristics. Both species are perennial climbing vines that form extensive patches and overgrow and smother native vegetation.
Black swallow-wort intertwined on itself. Photo: Leslie J. Mehrhoff, University of Connecticut, Bugwood.org
a stand of black swallow-wort. Photo: Leslie J. Mehrhoff, University of Connecticut, Bugwood.org
A stand of pale swallow-wort. Photo: Leslie J. Mehrhoff, University of Connecticut, Bugwood.org
Pale swallow-wort intertwined on itself with seed pods present. Photo: Chris Evans, Illinois Wildlife Action Plan, Bugwood.org
Origin
Black swallow-wort is native to the western Mediterranean region. Pale swallow-wort is native to Ukraine and southeastern European Russia.
Habitat
Black and pale swallow-wort tolerate a variety of soils and environmental conditions. They are typically found in upland habitats and have been observed in rocky shores, agricultural and natural fields, and woodlands. Swallow-wort favors full sun, but may thrive in semi-shaded areas as well. Pale swallow-wort also can be found in densely shaded sites. These species are disturbance- and drought-tolerant and can quickly spread once established.
Introduction and Spread
Swallow-wort may have been originally brought to North America for horticultural purposes as an ornamental plant. The shoots of swallow-wort start to emerge from the soil in the spring. The plants flower in the early summer and produce seed pods towards late July and early August. A square meter stand of swallowwort is capable of producing 1000-2000 seeds per year, which are then dispersed by wind. Black and pale swallow-wort are capable of self-fertilization and are polyembryonic, meaning each seed may contain multiple seedling producing embryos (Lumer and Yost 1995; St. Denis and Cappuccino 2004). The main method of spread is by seed; however, if the main stem is damaged, the perennating buds on the root crown will activate to produce axillary tillers (Weston et al. 2005).
Black swallow-wort roots. Photo: Leslie J. Mehrhoff, University of Connecticut, Bugwood.org
Black swallow-wort seeds. Photo: Leslie J. Mehrhoff, University of Connecticut, Bugwood.org
Black swallow-wort seed pods. Photo: Leslie J. Mehrhoff, University of Connecticut, Bugwood.org
Pale swallow-wort seed pods and seeds. Photo: John M. Randall, The Nature Conservancy, Bugwood.org
Impacts
Black and pale swallow-wort are moderately aggressive. The extensive patches of swallow-wort may overgrow native plants and disrupt natural succession. Herbivores and grazers, such as deer, tend to avoid swallow-wort to feed on more palatable plants. Additionally, due to the tangling nature of swallow-wort, large patches can be difficult to walk through and may interfere with forest management and recreation.
Lower arthropod diversity and abundance have been observed on invasive pale swallow-wort when compared to native plants (Ernst and Cappuccino 2005). Bird populations and soil microbial communities may also be indirectly impacted when swallow-wort invades. Pale swallow-wort grows particularly well on limestone-based soils and is invading globally rare alvar habitats in New York (DiTommaso et al. 2005).
Studies have looked at the potential impact of swallow-wort on monarch butterflies (Danaus plexippus). Most monarch butterflies tend not to lay their eggs on the two swallow-wort species, but if they do the larvae cannot survive. Additionally, the butterflies may indirectly be threatened if swallow-wort outcompetes their preferred larval host, common milkweed (Asclepias syriaca) (DiTommaso and Losey 2003).
Identification
Pale and black swallow-wort look very similar when not in flower. Both species have long, oval, opposite leaves (about 3-4 inches long by 2-3 inches wide). The leaves of pale swallow-wort tend to be a slightly lighter shade of green than black swallow-wort. Flowers of both species are small and star-like. Pale swallow-wort has a pink to maroon flower with petals that have a narrow base and no hairs, whereas black swallow-wort’s are a deep purple-black and have petals with a wider base (about half as wide as they are long) and hairy inner petals. Both species produce seed pods that are 1.5″ to 3″ long with wind-distributed seeds inside. Swallow-worts possess a fibrous root system.
Flowers of black swallow-wort. Photo: Leslie J. Mehrhoff, University of Connecticut, Bugwood.org
Flowers of pale swallow-wort. Photo: Leslie J. Mehrhoff, University of Connecticut, Bugwood.org
Flowers and foliage of pale swallow-wort. Photo: Leslie J. Mehrhoff, University of Connecticut, Bugwood.orgLine drawing of black swallow-wort (Pale swallow-wort would look similar). Illustration: USDA PLANTS Database, USDA NRCS PLANTS Database, Bugwood.org
Prevention and Control
Mechanical and manual control:
Mechanical and manual control of swallow-wort are difficult due to the presence of a deep, fibrous root system. If you dig up either of these plants, try to get as much of the root crown as possible. For small stands, the plants and seed pods can be collected in plastic bags and destroyed or disposed of properly in a landfill before they release their seed. Collecting seed pods is not a long-term solution and may be impractical for large stands.
Mowing will not eradicate the plant, but can be utilized to prevent seed formation if timed correctly. If swallow-wort stands are cut too early, the plants can recover and still produce viable seed. It is suggested to mow when immature seed pods are just starting to develop and do not yet contain viable seeds (McKague and Cappuccino 2005). Mowing of swallow-wort after seed development is not recommended as it could aid in seed dispersal.
Early seed pods starting to develop on pale swallow-wort. Photo: Rob Routledge, Sault College, Bugwood.org
Chemical Control:
Chemical herbicides, such as glyphosate or triclopyr, may be used for control as foliar sprays or on cut stems (Weston et al. 2005). Recent work in central New York State has shown that applying these herbicides in late summer (i.e. mid- to late-August) to plants that have been mowed or trimmed in early July may provides the best results. The mowing of plants in early July also ensures that no viable seeds are produced prior to application of the herbicides (DiTommaso et al., unpublished). These chemicals work on living plants and need ample leaf surface area to absorb the herbicide to effectively kill the roots. For this reason, it is not recommended to apply them on seedlings soon after emergence. Results may been seen in one to two weeks, but repeated application may be needed. Check with your local extension office about herbicide use and permits needed for chemical application. As always, with any chemical, follow the instructions on the label.
Biological Control:
At this time, there are no approved biological control methods for swallow-wort. Biological controls for swallow-wort species are currently being investigated.
Occurrences
There have been reports of invasive black swallow-wort from Ontario, Canada south to Pennsylvania and from the northern Atlantic coast west to Missouri. Pale swallow-wort is most abundant in the lower Great Lakes Basin and has been observed in New York, Connecticut, Indiana, Massachusetts, Michigan, New Hampshire, New Jersey, and Pennsylvania as well as in parts of Canada.
New York Distribution Map
This map shows confirmed observations (green points) submitted to the NYS Invasive Species Database. Absence of data does not necessarily mean absence of the species at that site, but that it has not been reported there. For more information, please visit iMapInvasives.
Female Sirex woodwasp Gyorgy Csoka, Hungary Forest Research Institute, Bugwood.org
Sirex Woodwasp, Sirex noctilio, larvae are woodborers that tunnel deep into the trunks of all species of pine (Pinus) trees. As the female Sirex woodwasp, Sirex noctilio, prepares to lay her eggs, she secrets a toxic mucus and symbiotic fungus into the tree which work synergistically to weaken, and in some cases kill the attacked tree(s). The mucus creates a suitable environment for the fungus which decays the wood so it is easier for the larva to digest.
Sirex woodwasp has caused mortality of millions of North American pines planted In Southern Hemisphere forests, where Sirex Woodwasp (S. noctilio) is also an invasive species. Currently, Sirex woodwasp is estimated to cause between $16 and $60 million in annual damages per year in those forests.
In the US, southern and western native pine forests and plantations face both an economic and ecological threat with the potential loss of valuable pine stands. The USDA Forest Service estimates the Sirex woodwasp could cost between $2.8 and $17 billion in lost sawtimber and pulpwood values if it becomes a nationwide established species.
History and Range
The Sirex woodwasp, S. noctilio, is native to Europe, Asia and north Africa. It was first discovered in North America in 2004 in a trap in Fulton, NY (Oswego County). In its native habitat it is considered a secondary or minor pest. Where it has been introduced it is considered to be a major pest. In its native range S. noctilio attacks Scots (P. sylvestris) Austrian (P. nigra) and maritime (P. pinaster) pines and is known to attack North American pines including red (P. resinosa), loblolly (P. taeda), slash (P. ellotti), ponderosa (P. ponderosa), lodgepole (P. contorta) and Monterey (P. radiata). White pine (P. strobus) is also attacked but is less preferred.
Pines 6” and larger are susceptible, stressed, suppressed and crowded pines seem to be favored. It is thought that the Sirex woodwasp can spread approximately 25 miles per year.
Sirex woodwasp known range, as of 2011. USDA/APHIS/PPQWorld Distribution of Sirex noctilio, 2000. Dennis Haugen, Bugwood.org
Identification
Female and Male Sirex Woodwasp Vicky Klasmer, Instituto Nacional de Tecnologia Agropecuaria, Bugwood.org
Adult Sirex woodwasp’s head and thorax are blue-black and have black antennae. Females have a blue-black abdomen with orange legs, while the abdomen of males has an orange mid-section and they have black hind legs. Woodwasps are 1 to 1.5 inches long and lack the typical narrow “waist” that most other wasps and hymenoptera have and there is a pointed plate that projects from the end of the abdomen.
There are many native woodwasps (Siricid woodwasps), and it is recommended to have a suspect woodwasp identified by an expert. Our native woodwasps only feed on dead and dying trees, while the Sirex woodwasp can also attack healthy trees. Images and descriptions of many Siricid woodwasps can be found in the Canadian Journal of Arthropod Identification “Siricidae (Hymenoptera: Symphyta: Siricoidea) of the Western Hemisphere” at http://www.biology.ualberta.ca/bsc/ejournal/sgsbws_21/sgsbws_21.html.
Larvae are 0.04 to 1 inch long, are creamy white and legless and have a spine at the end of their abdomen which is diagnostic for all woodwasps (also called horntails).
Larva – Vicky Klasmer, Instituto Nacional de Tecnologia Agropecuaria, Bugwood.org
Biology
The female Sirex woodwasp injects her ovipositor into the trunk of a pine tree, just into the wood, and injects one egg at a time along with the symbiotic fungus, Amylostereum areolatum, and a toxic mucus. The toxic mucus suppresses the tree’s defenses and decreases the tree’s ability to protect itself from the wood decaying fungus. When the larva hatches, it feeds on the fungus decayed wood and enzymes, and possibly the fungus as well, as it bores through the wood. Females lay 25 to 400 eggs. Fertilized eggs become females and unfertilized eggs become males. Larvae feed in the tree for 10-11 months and then pupate near the bark. Adults emerge after three weeks. Peak adult emergence is in July, but can occur from July through September. There is believed to be one generation per year in New York.
Many of the specifics of Sirex woodwasp biology and the relationship between Sirex and the fungus are still unknown. Research is currently being conducted that will hopefully provide more detail on this fascinating insect and its associated toxic mucus and symbiotic fungus.
Signs and Symptoms
Infested trees initially wilt, needles droop, then change from green to light green to red. This usually takes 3-6 months. Wilted trees generally die within a year.
When adults emerge they bore a 1/8 to 3/8 inch round exit hole through the bark. Trees in newly infested areas have exit holes 10-30 feet above ground. In heavily infested areas, the whole trunk is utilized by the larvae and exit holes can be seen along its length. There may be lines of pitch or resin beads at oviposition (egg laying) sites.
In the wood, frass packed larval galleries tunnel in towards the heartwood and then loop back out towards the bark. The fungus causes a brownish stain in the outer sapwood.
Wilted needles – Dennis Haugen, Bugwood.orgExit holes – Dennis Haugen, Bugwood.orgPitch from oviposition sites – Dennis Haugen, Bugwood.orgStaining caused by Amylosetereum fungus – Dennis Haugen, Bugwood.org
Damage in New York
Damage and galleries – Vicky Klasmer, Instituto Nacional de Tecnologia Agropecuaria, Bugwood.org
In New York State the largest damage is being seen in plantation Scots, Austrian, and red pine. These plantations were planted in the early to mid-20th century and were often unmanaged and are now crowded, stressed and underperforming. The death of these underperforming trees is not having a large economic or environmental effect in New York.
Damage in the rest of the US
Whereas Sirex woodwasp damage in New York is not profound, Sirex Woodwasp damage has the potential to be much greater in the South and Pacific Northwest. The projected economic damage to these forests is upwards of $17 billion dollars with hundreds of thousands of trees lost.
Biological control using a parasitic nematode, Deladenus siricidicola, has been successful in some infestations. The nematode infects the woodwasp larvae and ultimately sterilizes the females. These females lay infertile eggs that are instead filled with nematodes. As nematode populations grow, they can keep Sirex Woodwasp populations below damaging levels. Easy to rear in the lab, Deladenus siricidicola can be injected into infested trees. For more information about the lifecycle of Deladenus siricidicola see Cornell University’s Biological Control website:https://biocontrol.entomology.cornell.edu/pathogens/Deladenus.php
There are also parasitoid wasps, many native to North American, which could assist in managing Sirex woodwasp populations.
New York Distribution Map
This map shows confirmed observations (green points) submitted to the NYS Invasive Species Database. Absence of data does not necessarily mean absence of the species at that site, but that it has not been reported there. For more information, please visit iMapInvasives.
Dying Oak Wilt Infested Trees – William M. Ciesla, Forest Health Management International, Bugwood.org
Oak wilt (Ceratocystis fagacearum) is a systemic, lethal disease caused by a fungus. The fungus grows in the xylem (sapwood) of the infected oak eventually resulting in the vessels clogging with gums and tyloses and death of vascular cells which then prevents the uptake and movement of water.
In areas where oak wilt is established, the disease causes up to an 11% loss of annual growth in oak. (1)
Currently, there are few known infestations of Oak Wilt in New York State. These locations include Islip, Riverhead and Southhold in Suffolk County; Brooklyn in Kings County and Canandaigua in Ontario County. Oakwilt is also found in Glenville, Schenectady County. A small infestation was discovered and treated in 2008 but was found again in 2013. See below for more details on that outbreak. If you think your oak tree is infected with Oak Wilt, please contact your local Cornell Cooperative Extension or NYS Department of Environmental Conservation Lands and Forests office immediately. [Information updated as of 2018]
Hosts
Red oak is most severely affected and white oak is moderately affected. An infested tree begins to show symptoms in early summer. Red oaks typically perish within two month whereas white oaks can survive for many years with the disease.
Biology and Identification
Foliar symptoms of oak wilt in red oak. – Fred Baker, Utah State University, Bugwood.org
Ceratocystis fagacearum, the oak wilt fungus, is spread to an oak tree via root grafts or beetles feeding on sap (nitidulids) at open wounds or on the leaves of healthy trees (oak bark beetles). Once inside the tree, the oak wilt fungus begins to replicate within the xylem of the tree.
Symptoms first appear near the top of the canopy. The outside of the leaves turn bronze, brown, or dull green, usually starting at the top of the leaf, while the base of the leaf remains green. Some leaves curl, droop and wilt. Leaves begin to drop soon after symptoms first develop.
In some species of oak, brown streaks develop in the outer sapwood of infected branches when leaf symptoms appear. This can be seen when an infected branch is cut diagonally. Eventually the tree loses all its leaves to the wilt.
Brown streaks visible in oak branches. – T.W. Bretz, USDA Forest Service, Bugwood.org
The year following the defoliation, fungus or spore mats may form on the trunk under the bark and on the opposing wood. Oval mats are gray or tan and turn black with age. The mats develop pressure pads that push out and crack the bark, exposing the fungus mat. The mat has a fruity odor which attracts then contaminates sap feeding beetles. Those contaminated beetles then carry the fungus to open wounds on other oak trees. Mats are primarily formed on red oaks.
Fungus Mat growing on the wood of an infected oak tree. – T.W. Bretz, USDA Forest Service, Bugwood.org
Spread
Oak wilt is primarily spread by beetles which feed on the fungus mats under the bark then carry spores to open wounds on nearby oak trees as well as through underground root grafts between oak trees. Infected trees form infection centers and mortality of oak occurs in rings from the initial infestation as the fungus moves through naturally occurring root grafts.
Root grafted oak trees usually die 1-6 years after the first oak is infected, and disease centers can move up to 50 feet per year.
Oak wilt can also be spread long distances through the movement of infested firewood.
Oak Root Grafts – Joseph O’Brien, USDA Forest Service, Bugwood.org
Oak Wilt Disease Center – Joseph O’Brien, USDA Forest Service, Bugwood.org
Control and Prevention
Vibratory Plow – Joseph O’Brien, USDA Forest Service, Bugwood.org
If an oak tree in a known oak wilt infested area needs to be pruned, do not prune in the spring or summer, especially April, May and June when the fungal mats and the sap beetles are present. If it is necessary to prune, the use of wound paint will prevent contaminated beetles from spreading the fungus to the open wound. (Note: This is one of the few times wound dressing is recommended; normally the pruning wound should be allowed to heal exposed to the air.) Clean pruning equipment between cuts and trees. Do not use climbing irons/spikes on living oak trees.
Vibratory plows or backhoes can be used to break root grafts by creating a trench around individual or groups of trees. This severs roots which prevents the spread through the grafts.
Trench dug to break root grafts. – NYS Dept of Environmental Conservation
Individual oak trees can also be treated with systemic fungicides. (Only if disease becomes more prevalant in NY. Until then, the tree should be removed.)
If you think you have an infected tree contact your local Cornell Cooperative Extension or DEC Lands and Forest office immediately. Trees found to be infected in New York should be cut, chipped, and burned.
In 2008, a small infestation (76 trees) in Glenville, NY (Schenectady County) was found by an observant homeowner. He called his local Cornell Cooperative Extension horticulture educator who recognized it as oak wilt. Once the disease was confirmed by the experts at the USDA Forest Service, the NYS Department of Environmental Conservation –Forest Health Unit was able to quickly mobilize to survey and respond. All infected trees were removed and root zones were trenched (to sever root grafts). Monitoring has continued for three years and no additional oak wilt has been found. If no oak wilt is found in 2012 it will be deemed a successful eradication.
This case is a superb example of interagency cooperation between NYS DEC, NYS Agriculture and Markets, APHIS, Cornell University and Cooperative Extension, and local officials.
Aerial Photo of dead oak trees. – NYS Dept of Envirnmental ConservationNYS Oak Wilt Infection Area- Before – NYS Dept of Envirnmental ConservationNYS Oak Wilt Infection Area – After – NYS Dept of Environmental Conservation
One of New York’s most striking and dangerous invasive plants, the giant hogweed (Heracleum mantegazzianum) can make a case of poison ivy seem like a mild rash.
History and Introduction
A member of the carrot and parsley family of plants (Apiaceae), giant hogweed is native to the Caucasus region of Eurasia. Because of its unique size and impressive flower head, the plant was originally introduced to Great Britain as an ornamental curiosity in the 19th century. The plant is named after the mythological god Hercules (he of robust size and strength). It was later transported to the United States and Canada as a showpiece in arboreta and Victorian gardens (one of the first North American plantings of giant hogweed was in gardens near Highland Park in the City of Rochester, New York). It was also a favorite of beekeepers because of the size of its flower heads (the amount of food for bees is substantial). A powder made from the dried seeds is also used as a spice in Iranian cooking. Unfortunately, as with so many invasive plants, giant hogweed escaped cultivation and has now become established in a number of areas in New York (see map, below), as well as in Connecticut, the District of Columbia, Illinois, Maine, Maryland, Massachusetts, Michigan, Ohio, Oregon, Pennsylvania, Washington, Wisconsin, and Ontario and Vancouver Island in Canada. Because of its public health hazard potential and, to a lesser extent, to its potential ecological impacts, giant hogweed is on the federal noxious weed list and several state lists of prohibited plant species.
Density of Observations, Giant Hogweed. iMapInvasives.org
Biology & Habitat
Mature giant hogweed plants
Giant hogweed (Heracleum mantegazzianum) is a member of the carrot or parsley family, Apiaceae (Umbelliferae). Except for its size, the plant can be mistaken for a number of native, noninvasive plants such as cow parsnip (Heracleum lanatum), Angelica (Angelica atropurpurea), and poison hemlock (Conium maculatum). Of these, the plant most likely to be misidentified as giant hogweed is cow parsnip. A fourth, not-so-innocuous, invasive giant-hogweed imposter found throughout North America is wild parsnip (Pastinaca sativa). Information on how to distinguish these giant hogweed wannabees from the real thing can be found later in this profile.
Giant hogweed is a perennial herb with tuberous root stalks. It survives from one growing season to another by forming perennating buds (surviving from season to season) and enduring a period of dormancy during the winter. The plant develops numerous white flowers that form a flat-topped, umbrella-shaped head up to two and a half feet across, resembling “Queen Anne’s Lace on steroids.” Flowers form from late-spring through mid-summer. Numerous (up to 100,000), half inch long, winged, flattened oval seeds form in late-summer. These seeds, originally green, turn brown as they dry and can be spread by animals, surface runoff of rain, or on the wind, establishing new colonies. Seeds can remain viable in the soil for up to 10 years. The plant’s stems die in the fall and remain standing through the winter, topped with the huge, brown dead flower heads.
Giant hogweed’s thick hollow stems are generally one to three inches in diameter but can reach four inches. Also impressive are the plant’s lobed, deeply incised compound leaves, which can reach up to five feet in width. The plant may grow to 15 to 20 feet in height.
Giant hogweed leaves
Identification
As mentioned earlier, there are several plants in New York and the Northeast that can be mistaken for giant hogweed. Key features for distinguishing these plants from giant hogweed are explained below. Click the identification tables to enlarge.
Giant hogweed may grow to 15 to 20 feet in height. Stems are 1 to 3 inches in diameter, but may reach 4 inches. Stems are marked with dark purplish blotches and raised nodules. Leaf stalks are spotted, hollow, and covered with sturdy bristles (most prominent at the base of the stalk). Stems are also covered with hairs but not as prominently as the leaf stalks. Leaves are compound, lobed, and deeply incised; can reach up to 5 feet in width. Numerous white flowers form a flat-topped, umbrella-shaped head up to two and a half feet across. [See ID table 1]
Native Cow parsnip, while resembling giant hogweed, grows to only five to eight feet tall. The deeply ridged stems can be green or slightly purple, do not exhibit the dark purplish blotches and raised nodules of hogweed, and only reach one to two inches in diameter, contrasted with hogweed stems which can reach three to four inches in diameter. Where giant hogweed has coarse bristly hairs on its stems and stalks, cow parsnip is covered with finer hairs that give the plant a fuzzy appearance. Both sides of the leaves exhibit these hairs but they are predominantly on the underside of the leaves. In contrast to hogweed’s two- to two-and-a-half-foot flower heads, cow parsnip flower clusters are less than a foot across. The size difference carries over into leaf size with hogweed’s five foot, deeply incised leaves replaced by leaves that are less incised and only two- to two-and-a-half-foot across.
Native purple-stemmed Angelica is more easily differentiated from giant hogweed by its smooth, waxy green to purple stems (no bristles, no nodules), and its softball-sized clusters of greenish-white or white flowers, seldom reaching a foot across. As with cow parsnip, Angelica is much shorter than giant hogweed, usually no more than eight feet tall. Angelica leaves are comprised of many small leaflets and seldom reach more than two feet across.
Poison hemlock, a non-native biennial, is also shorter than giant hogweed, growing to only four to nine feet in height. While the stem has some purple blotches, it is waxy and the entire plant (stems, stalks, leaves) is smooth and hairless. The leaves are dramatically different from those of hogweed, being fernlike and a bright, almost glossy, green. All branches have small flat-topped clusters of small white flowers. Another distinguishing characteristic is poison hemlock’s unpleasant mouse-like odor. The entire plant is toxic, and the volatile alkaloids can even be toxic when inhaled. [See ID Table 2]
Wild parsnip, like giant hogweed, is of special concern because it, too, can cause phytophotodermititis, only not usually as severe as that of giant hogweed. This plant can be found extensively throughout NY’s Southern Tier, in the region east of Lake Ontario, some Central and Western New York counties, parts of the Catskills and counties east of the Hudson River. Unlike the perennial giant hogweed, wild parsnip is biennial, producing a rosette of leaves close to the ground in its first year and a single flower stalk with a flat-topped umbel with clusters of yellow flowers in its second year. The plant reproduces by means of the seeds of these flowers; it does not re-grow from its root as does giant hogweed. Wild parsnip is much smaller than giant hogweed, seldom exceeding 5 feet in height. Wild parsnip stems are yellowish-green with vertical grooves running their length. Wild parsnip has compound pinnate leaves with 5 to 15 toothed and variably lobed yellowish-green leaves.
Ecological Impacts
Colonies of giant hogweed can become quite dense owing to the plant’s prolific seed production and rapid growth rate. Such dense stands crowd out slower growing plants, the thick hogweed canopy displacing native plants that need direct sunlight to grow. The decreased abundance of beneficial native plants can reduce the utility of the area for wildlife habitat. When riparian plants are displaced, stream bank erosion can increase and streambeds can become covered with silt.
Human Health Impacts
Giant hogweed phytophotodermatitis
Giant hogweed is one of a very few North American invasive plants that can cause human health impacts as well as ecological damage, causing a significant reaction when humans come in direct contact with the plant. Spread of this plant in urban and suburban areas is viewed as an incipient public health hazard. [Wild parsnip can result in almost as severe reactions.]
Soon after humans bruise the leaves or stems of the more common poison ivy, poison oak, and poison sumac, an allergic reaction to the plants’ poisonous oil (akin to carbolic acid) causes significant skin irritation, itching, rashes and open sores. In the case of giant hogweed, however, simply simply brushing against the plant’s leaves or stems does not cause the skin inflammation. For giant hogweed to affect a person, sap from a broken stem or crushed leaf, root, flower or seed must come into contact with moist skin (perspiration will suffice) with the skin then being exposed to sunlight. Irritation is not immediate, but will usually appear within one to three days after exposure. This form of skin irritation (dermatitis) is called “phytophotodermatitis”. The plant’s clear, watery sap contains a glucoside called furanocoumarin that is a psoralen. Psoralens sensitize the skin to ultraviolet radiation and can result in severe burns, blistering, painful sores, and purplish or blackened scars. These phototoxic effects are the result of the binding of the psoralens to nuclear DNA under the influence of ultraviolet irradiation, and the subsequent death of affected cells.
The first signs of giant hogweed-caused photodermatitis are when the skin turns red and starts itching. Within 24 hours, burn-like lesions form, followed by large, fluid filled blisters within 48 hours. The initial irritation usually will subside within a few days, but affected areas may remain hypersensitive to ultraviolet light for many years and re-eruptions of lesions and blisters may occur. On rare occasions, particularly in very sensitive individuals, the burns and blisters may be bad enough to require hospitalization. A side effect of exposure to the psoralens is the production of excessive amounts of melanin in the skin, resulting in residual brown blotches called hyper-pigmentation; scars and brown to black blotches may last for several years. The worst risk of exposure to giant hogweed is to one’s eyes – getting even minute amounts of the sap in the eyes can result in temporary or even permanent blindness. Medical help should be sought immediately; by the time symptoms of burning and hypersensitivity to sunlight are apparent, the damage could already be irreversible.
The only known antidote to contact with the sap is to immediately wash skin thoroughly with soap and water, removing the sap and hopefully preventing any reaction with subsequent exposure to sunlight. Once the irritation begins, medical advice should be sought. Treatment with prescription topical steroids early on may reduce the severity of a person’s reaction. It will also be important to cover the burns and blisters with light sterile dressings to prevent infection. Long-term, use of sunblock in subsequent years may be required to prevent sensitization by sunlight again.
People most at risk include landscape technicians and yard maintenance laborers who may come in contact with the sap when cutting the plant down or using line trimmers to control new growth. Children breaking off the long, bamboo-like stems to use as play swords are also at great risk. However, sometimes direct contact with the plant is not necessary for a reaction. Farmers have been known to develop symptoms when they touch cows who have gotten the sap on their skin while grazing (cows, themselves, seem impervious to the sap). The best prevention measure is to wear long sleeves and long-legged pants when contact with the plant is a possibility.
Control
If it weren’t for giant hogweed’s public health impacts, the plant most likely would not be worth the effort of controlling it. Although it does have ecologic impacts, they are not as severe as many other wetland invasive plants. However, the health impacts can be severe and the plant has found itself on the federal noxious weed list and several state lists, as well. It is a particular target of parks and transportation/highway departments’ invasive plant eradication efforts. Such eradication programs can incorporate a combination of physical removal and chemical control. If undertaken properly, such programs can be done without harm to humans or damage to the environment. Recently some landowners have been known to refuse permission to allow highway departments to chemically treat giant hogweed thickets. It is believed that this is usually a case of lack of knowledge on the landowner’s part.
Giant hogweed is very difficult to eradicate. Although the stems, stalks, leaves and flowers can be killed with a number of common selective herbicides, such as 2,4-D (the third most-often used herbicide in North America), dicamba (a benzoic acid herbicide), TBA (terbuthylazine) and MCPA, these herbicides are not effective at killing the plant’s tuberous perennial roots. Another common, selective broadleaf herbicide, triclopyr (a common brand name is Brush-B-Gone®), is also effective, particularly when applied directly to the entire surface of leaves and stems during periods of active growth; numerous applications may be needed to kill the root stalk. Early application (during the bud stage and the period of active plant growth) of glyphosate (commonly sold under the trade names Rodeo® and Roundup®) is most effective. Care should be taken when using any herbicides to control giant hogweed; particular care should be taken when using glyphosate as it is nonselective and will kill both the hogweed and desirable plants such as grass. Before using any herbicide, check with your state environmental regulatory agency to find out which products are licensed for homeowner use in your state.
For those hesitant to utilize herbicides, giant hogweed can be managed using various “cultural” methods. Unfortunately, owing to the plant’s persistence and spread by blowing seeds, such control can take many seasons worth of effort to achieve 100% control. Individual plants can be dug out, removing the entire rootstalk, a difficult process, particularly in patches where the plant has spread by root growth. Mowing, cutting and use of line trimmers can be used to remove a standing crop and starve the rootstalk. Unfortunately, unless performed numerous times during a season, mowing only serves to stimulate budding on the rootstalk. All of these methods should be done with extreme care and only while wearing protective clothing and eye protection. Skin contact with soiled clothing should also be avoided. Biocontrol by grazing cows and pigs (which are apparently not affected by the plant’s sap) may also help to manage but not eliminate the plant. Care should be taken not to get sap on uncovered skin when touching livestock after the animals contact crushed or bruised hogweed.
Control of wild parsnip is less difficult than controlling giant hogweed because as a biennial, wild parsnip reproduces only from seed, not from its rootstock. This plant can be controlled by cutting the stem from the root below ground level with a shovel, spade or machete before the seed head matures.
If you find giant hogweed in New York State, you are encouraged to call New York State’s Department of Environmental Conservation (DEC) Giant Hogweed Hotline:
845-256-3111
Reports
2014 Giant Hogweed Annual Report
The DEC Forest Health giant hogweed crews and partner agencies (Oswego SWCD and four PRISMs (Partnerships for Regional Invasive Species Management) — APIPP, CRISP, Lower Hudson, and SLELO) had an extremely productive field season which resulted in the following achievements:
1,495 (93%) of the 1,613 active sites located throughout 47 counties in NY State were visited
501 sites previously infested (28% of all sites), no longer had any giant hogweed plants
239 of these sites have been monitored for 3 years with no plants found and we now consider them eradicated
516 sites had only 1-19 plants (34% of active sites). The sites are getting smaller in size
556 sites were treated with root-cutting- 22,255 plants controlled
551 sites were treated with herbicide- 397,000 plants sprayed
316 sites were treated with flower/seed head removal- 7,677 flower/seed heads removed
The hotline, based out of the New Paltz DEC office, received 2,491 calls and emails
Several species of honeysuckle found in NY are characterized as invasive, including: Morrow’s honeysuckle (Lonicera morrowii), Tatarian honeysuckle (Lonicera tatarica), Amur honeysuckle (Lonicera maackii), and Japanese honeysuckle (Lonicera japonica). To the non-botanist, native and invasive non-native honeysuckles appear very similar. One way to distinguish between native and invasive honeysuckles is by looking at the stems – native honeysuckles have solid stems while invasive honeysuckles have hollow stems.
Habitats
All four species are successful invaders of a similar range of habitats, including: abandoned fields; pastures; early successional, open canopy, and planted forests; along the edge of woodlots; floodplains; highway, railway and utility rights-of-way; open disturbed areas; vacant lots; edges of lawns; and, gardens. L. japonica can also be found in agricultural fields. All four grow best in full sun; L. japonica is the most shade-tolerant of the four, with L. tatarica and L. maackii being semi-shade tolerant.
Origin and Introduction
L. tatarica is native to Central Asia and Southern Russia and is believed to have been introduced into North America for ornamental purposes as early as the 1750s. L. japonica, – a native of China, Japan and Korea – was introduced for horticultural purposes in 1806 on Long Island; it was widely distributed as a garden plant through the early-1900s when it was finally recognized as a weed. L. maackii, also native to China, Japan and Korea, was introduced as seeds to arboreta throughout the U.S. in the late-1800s to determine whether the plant would grow in North America. This species of honeysuckle was utilized as a soil stabilization and wildlife planning until the mid-1980s and is still available for sale on-line. L. morrowii, a native of Japan, was imported to Massachusetts in the 1860s and was later released as an ornamental. All four species have escaped cultivation and are easily spread by birds.
Identification
L. morrowii, L. tatarica, and L. maackii), are perennial shrubs; L. japonica is a perennial woody vine (although its leaves can remain green throughout mild winters). The shrub forms range from 6 to 15 feet in height, while vines can reach 30 feet in length. The egg-shaped leaves range from 1 to 3 inches in length and are arranged oppositely along stems. Invasive honeysuckles begin flowering from May to June and bear small (less than 1 inch long), very fragrant tubular flowers ranging from creamy white through various shades of pink to crimson. L. morrowii and L. tatarica produce ¼ inch red berries from mid-summer through early-fall; L. maackii’s dark-red berries don’t ripen until late-fall; L. japonica produces dark-purple or black berries in the fall. Stems of all four are hollow.
Impacts
All three species can form very dense populations that can outcompete and suppress the growth of native plant species. These dense stands suppress the growth of other native species. L. maackii leafs out very early in spring, giving it a competitive advantage over native plants. L. japonica leaves are semi-evergreen allowing the plant to grow longer into the winter and giving it a competitive advantage over native vegetation. It shades out understory growth preventing the success of native understory plants and tree seedlings. Its vigorous vine growth covers native trees; the weight of the vine growth can bring down weak trees. By decreasing light availability to the understory, these invasive honeysuckles can alter habitats by depleting soil moisture and nutrients. The invasive honeysuckle berries do not contain the amount of fat and nutrients present in native honeysuckle berries; eating large amounts of the less nutritious invasive berries rather than native berries can have negative impacts on migrating.
Prevention and Control
Because these plants spread rapidly via birds eating seeds, control should be started in late-summer or early-fall before seeds are ready to be dispersed. In early stages of invasion, or in cases where populations are at low levels, hand removal of honeysuckle seedlings or young plants is a viable option when repeated annually. Systemic herbicides can be utilized in cases of heavy infestation. Specific state rules should be followed and the appropriate (low environment impact, legally labeled for control of these plants) herbicides should be used. For invasive honeysuckles growing in open habitats, prescribed burning may be an effective control alternative.
New York Distribution Map
This map shows confirmed observations (green points) submitted to the NYS Invasive Species Database. Absence of data does not necessarily mean absence of the species at that site, but that it has not been reported there. For more information, please visit iMapInvasives.
A thicket of Japanese knotweed. Citation: Tom Heutte, USDA Forest Service, www.invasives.org
Background
Japanese knotweed (Polygonum cuspidatum Sieb. & Zucc.), a member of the buckwheat family, was introduced into the U.S. from Eastern Asia (Japan, China, Korea) as an ornamental on estates in the late-1800s. It has also been used as an erosion control plant. By the mid-1890s, it was reported near Philadelphia, PA, Schenectady, NY, and in New Jersey. Although once sold through seed and plant catalogs, by the late-1930s knotweed was already being viewed as a problematic pest. The plant, which can grow from three to 15 feet tall, has bamboo-like stems and is sometimes called Japanese bamboo. As with many invasive plants, knotweed thrives in disturbed areas and once established can spread rapidly, creating monoculture stands that threaten native plant communities. Japanese knotweed can tolerate deep shade, high temperatures, high soil salinity and drought. It is commonly found along streams and rivers, in low-lying areas, disturbed areas such as rights-of-way, and around old home and farmsteads.
Distribution
Japanese knotweed can be found in the U.S. in 42 states coast to coast except the arid Southwest, several of the deep South Gulf states and the highest of the Rocky Mountains. It is also in eight Canadian Provinces.
US Distribution of Japanese Knotweed. Map: USDA NRCS PLANTS Database
Identification
The plant is an upright, shrubby, herbaceous, woody-appearing perennial reaching heights of 10 to 15 feet (3 to 4.5 m). The stems are smooth, stout, and hollow. Where the leaves attach to the stem, the stem is swollen with a membranous sheath surrounding the joints. The leaves are broadly ovate (broad and rounded at the base and tapering toward the end), 3 to 6 inches (8 to 15 cm) long by 2 to 4 inches (5 to 10 cm) wide, alternating on stem, broadly oval to somewhat triangular or heart-shaped, pointed at the tip. The plant’s shoots come up from a network of spreading rhizomes. These horizontal roots can reach lengths of 65 feet (20 m) or more. Japanese knotweed has branched sprays of small greenish-white flowers from August to September. Although they have complementary male and female organs, those organs are vestigial and the flowers function unisexually. The plant develops small winged fruits Seeds: triangular, shiny, very small, about 1/10 inch (2.5 mm) long. Japanese knotweed spreads primarily by seed (transported by wind, water, animals, humans, or as a soil contaminant), stem fragments, and by shoots sprouting from its system of rhizomes.
Japanese knotweed showing oblate leaves and flowers. Jan Samanec, State Phytosanitary Administration, Bugwood.org
Jan Samanek, State Phytosanitary Administration, Bugwood.org – See more at: http://www.forestryimages.org/browse/detail.cfm?imgnum=5205100#sthash.WY0qNOzS.dpuf
Jan Samanek, State Phytosanitary Administration, Bugwood.org – See more at: http://www.forestryimages.org/browse/detail.cfm?imgnum=5205100#sthash.WY0qNOzS.dpuf
Jan Samanek, State Phytosanitary Administration, Bugwood.org – See more at: http://www.forestryimages.org/browse/detail.cfm?imgnum=5205100#sthash.WY0qNOzS.dpuf
Impacts
Japanese knotweed spreads rapidly, forming dense thickets that crowd and shade out native vegetation. This reduces species diversity, alters natural ecosystems, and negatively impacts wildlife habitat. The ground under knotweed thickets tends to have very little other growth. This bare soil is very susceptible to erosion, posing a particular threat to riparian areas. Once established, populations of Japanese knotweed are extremely persistent and hard to eradicate.
Prevention & Control
As with most invasive plants, Japanese knotweed can quickly inhabit disturbed ecosystems. One of the best ways to prevent its colonization is to ensure that disturbed habitats are rehabilitated with native vegetation before knotweed can invade.
Mechanical control
Single young plants can be pulled by hand depending on soil conditions and root development. If all of the root system isn’t removed, re-sprouting can occur. For small initial populations beyond single plants, or in environmentally sensitive areas where herbicides cannot be used, grubbing with a pulaski or similar tool to remove all of the roots after cutting back the standing vegetation can be an effective control measure. All parts of the removed plants should be bagged and disposed of in a secure location. It is best if knotweed not be mown or cut with weed trimmers as the pieces of the plant can easily get moved around and re-sprout, spreading, rather than controlling the plant.
Chemical control
Chemical controls for Japanese knotweed include application of glyphosate and triclopyr herbicides to freshly cut stems or to foliage. Before using chemicals to control any invasive plant, check with your local environmental or natural resources management agency or Cooperative Extension office to find out what chemicals are legal for use on knotweed in your area.
Catskill Regional Invasive Species Partnership (CRISP) Japanese knotweed video
New York Distribution Map
This map shows confirmed observations (green points) submitted to the NYS Invasive Species Database. Absence of data does not necessarily mean absence of the species at that site, but that it has not been reported there. For more information, please visit iMapInvasives.
The non-native Phragmites australis, or common reed, can rapidly form dense stands of stems which crowd out or shade native vegetation in inland and estuary wetland areas. Phragmites turns rich habitats into monocultures devoid of the diversity needed to support a thriving ecosystem. Non-native Phragmites can alter habitats by changing marsh hydrology; decreasing salinity in brackish wetlands; changing local topography; increasing fire potential; and outcompeting plants, both above and belowground. These habitat changes threaten the wildlife that depend on those wetland areas for survival.
Stand of Phragmites Credit: Bernd Blossey, Cornell University, Bugwood.org
History
Common reed, Phragmites australis, is in the Poaceae or grass family. There are at least three lineages, or strains, of common reed in the U.S. At least one is native to the U.S. including the one that was most common in New York, P. australis subsp. americanus. Another common reed strain, P. australis var. berlandieri may or may not be native to the U.S. and is found in California, along the Gulf Coast, and in the Southeast. One strain is non-native, and was accidentally introduced from Europe in the late 18th or early 19th century in ship ballast. This non-native strain is now the most common Phragmites found in New York and the Northeast. There is no field evidence that the non-native will hybridize with the native Phragmites at this time. This fact sheet focuses on the non-native Phragmites.
Biology
The non-native Phragmites is a perennial grass that can reach over 15 feet in height. It is often found in dense clonal stands made up of living stems and standing dead stems. Stems of the non-native Phragmites are hollow, usually green with yellow nodes during the growing season, and yellow when dry in the winter. Phragmites leaves are blue-green to yellow-green, up to 20 inches long and 1 to 1.5 inches wide at their widest point. They are arranged all along one side of a stem.
In late July and August, Phragmites is in bloom with purple to gold highly-branched panicles of flowers. The seeds are grayish and appear fluffy due to the silky hairs that cover each seed. Spread occurs through rhizomes, stolons, and seeds; stolons can grow up to 43 feet from the parent plant.
Phragmites in bloom Credit: Leslie J. Mehrhoff, University of Connecticut, Bugwood.org
Root growth belowground is also profuse. Phragmites forms a ticket of roots and rhizomes that can spread 10 or more feet and several feet deep in one growing season.
Each Phragmites plant produces thousands of seeds each year, but seed viability is low, although viability varies from year to year. New sites are established through seed movement and from rhizome fragments that float down stream or are moved in soil, especially along roadsides.
Large clumps of Phragmites can live for decades, but no part lives for more than 8 years.
There are physiological differences between the native Phragmites and the non-native Phragmites. See the Plant Conservation Alliance Phragmites Fact Sheet comparison table for details.
Phragmites in seed. Credit: Jil Swearingen, USDI National Park Service, Bugwood.org
Habitat
The non-native Phragmites occurs throughout the eastern half of the U.S. and in Colorado. In New York, Phragmites is ubiquitous, growing in roadside ditches and swales; tidal and non-tidal wetlands; freshwater and brackish marshes; river, lake and pond edges; and disturbed areas. It tolerates fresh and moderately-saline water and prefers full sun.
Credit: John M. Randall, The Nature Conservancy, Bugwood.org
Credit: John M. Randall, The Nature Conservancy, Bugwood.org
Management
Due to the similarity of non-native Phragmites and native Phragmites, proper identification of the grass is important before taking management action. Due to Phragmites growth in sensitive habitats, be sure to have a restoration plan in place for the area once Phragmites has been eliminated. Phragmites roots hold onto soil, and clonal colonies trap nutrients and organic matter and add to the organic matter in the soil. After Phragmites colonies are removed the soil may be more prone to erosion.
To control Phragmites a number of tactics may be used, but due to the many variables at each site many suggest that Phragmites management should be “site-specific, goal-specific, and value-driven.” Often multiple tactics are needed to ensure success. The best time to manage Phragmites is in midsummer when it is releasing pollen. Thorough monitoring and follow-up management are necessary to control shoots from surviving rhizomes.
Prevention
Maintain, or plant, vegetation that competes with Phragmites. Jesuit’s bark (Iva frutescens), groundsel-tree (Baccharis halimifolia), black rush (Juncus roemerianus), and saltmeadow cordgrass (Spartina patens) have been shown to limit Phragmites spread. Also, reducing nutrient loads may restrict the spread of Phragmites.
Mechanical
Repeated mowing may produce short-term results and repeated stem breakage in high-water years has been shown to kill large portions of Phragmites colonies. Hand pulling is not feasible due to the expansive and tough root and rhizome network. Root removal from the soil is not effective as small or broken portions of rhizomes left in the soil can create new plants.
Hydrologic
Manipulating the water level around Phragmites has been shown to decrease populations in some conditions. Consult the Element Stewardship Abstract for Phragmites australis produced by the Nature Conservancy for more information. http://www.invasive.org/gist/esadocs/documnts/phraaus.pdf
Chemical
There are herbicides available for Phragmites control. New colonies, with smaller root and rhizome systems, are easier to control with herbicides. Apply after the plant has flowered, in late summer or early fall. Applications can be foliar, cut stump, or injected. Multiple years of treatment may be necessary to eliminate any surviving rhizomes. Herbicides applied in wetland areas must be applied by a certified pesticide applicator. Contact your local Cornell Cooperative Extension office, http://www.cce.cornell.edu, for herbicide usage assistance. Always apply pesticides according to the label directions; it’s the law.
Fire
Prescribed burns have been shown effective when conditions are right, and can occur in conjunction with herbicides or water level management. To be successful as a stand-alone tool, burns need to be hot enough to kill rhizomes in the soil. After herbicide treatments, burns can remove standing dead stems to make way for desirable vegetation. Flooding after burns will limit soil air to surviving rhizomes. Burns should be conducted once flowering has occurred. For more information on controlled burns, see the USDA Forest Service Fire Effects Information System “Phragmites australis Fact Sheet,” Fire Effects section at http://www.fs.fed.us/database/feis/plants/graminoid/phraus/all.html#FIRE%20EFFECTS.
New York Distribution Map
This map shows confirmed observations (green points) submitted to the NYS Invasive Species Database. Absence of data does not necessarily mean absence of the species at that site, but that it has not been reported there. For more information, please visit iMapInvasives.






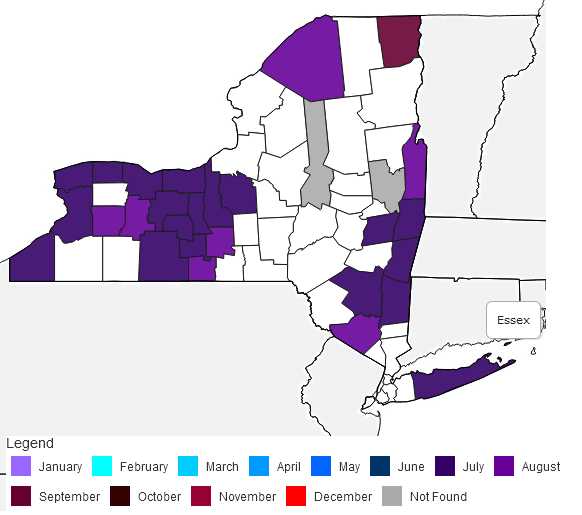

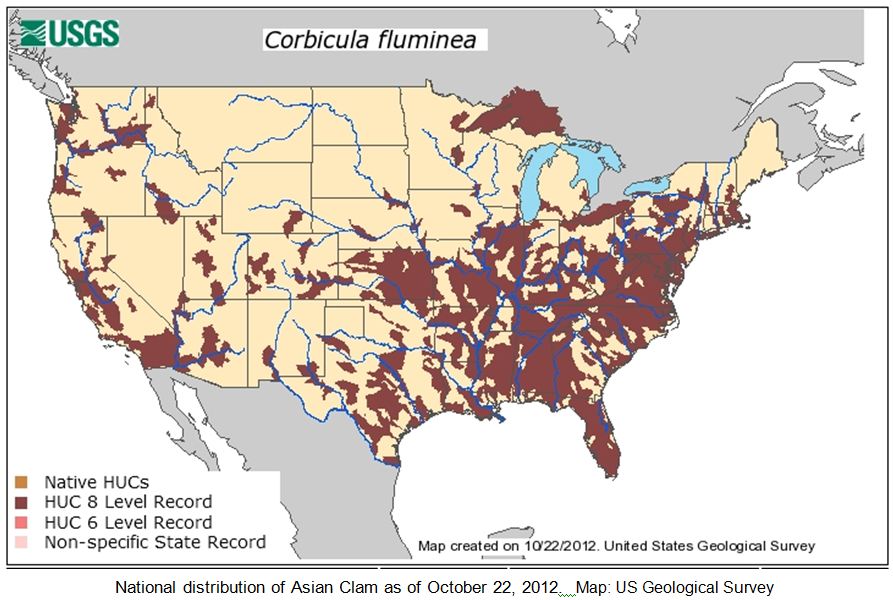
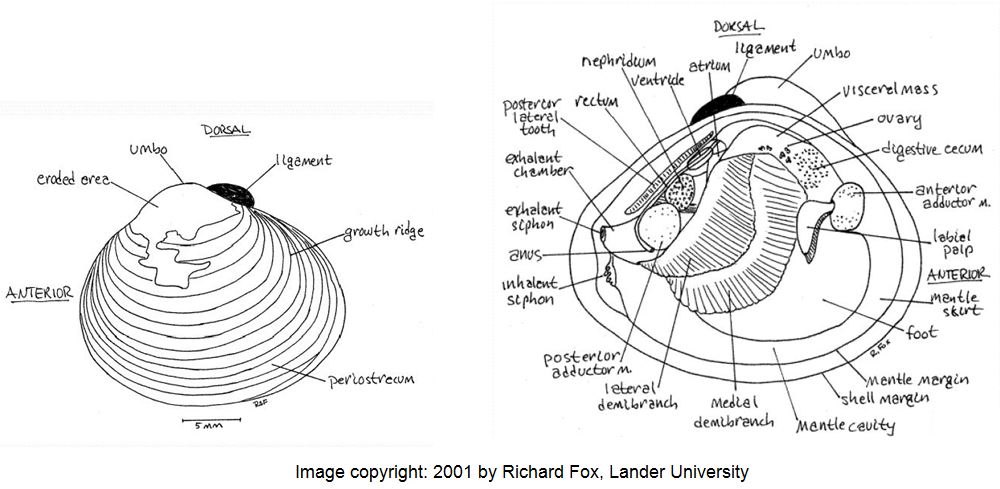
.JPG)

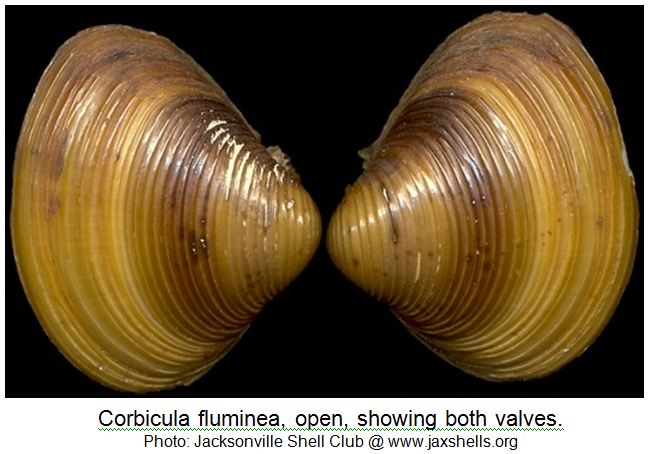
.jpg)













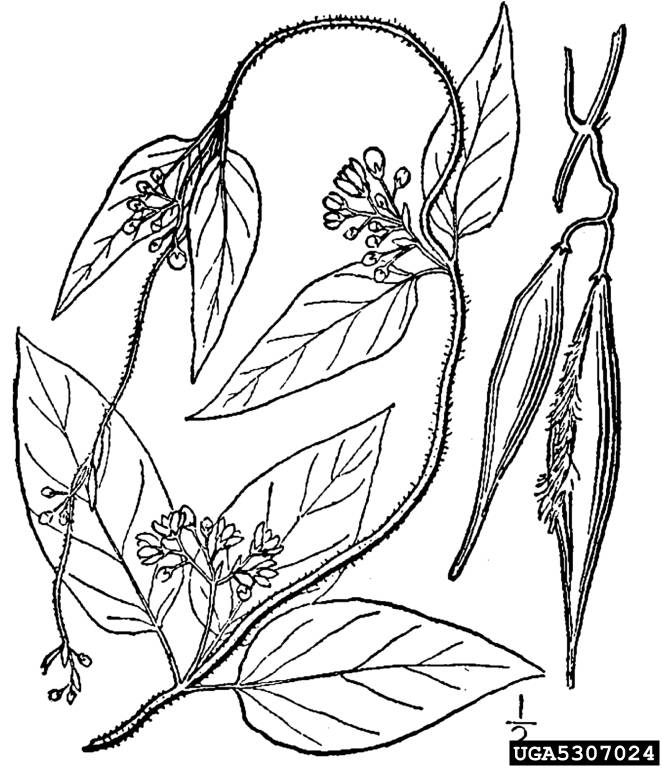


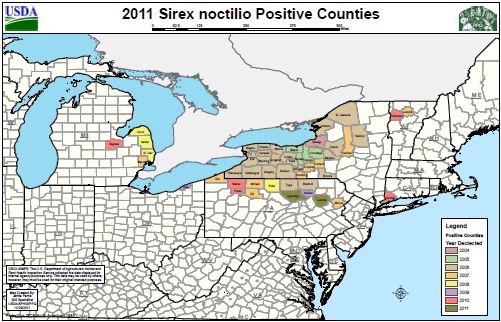
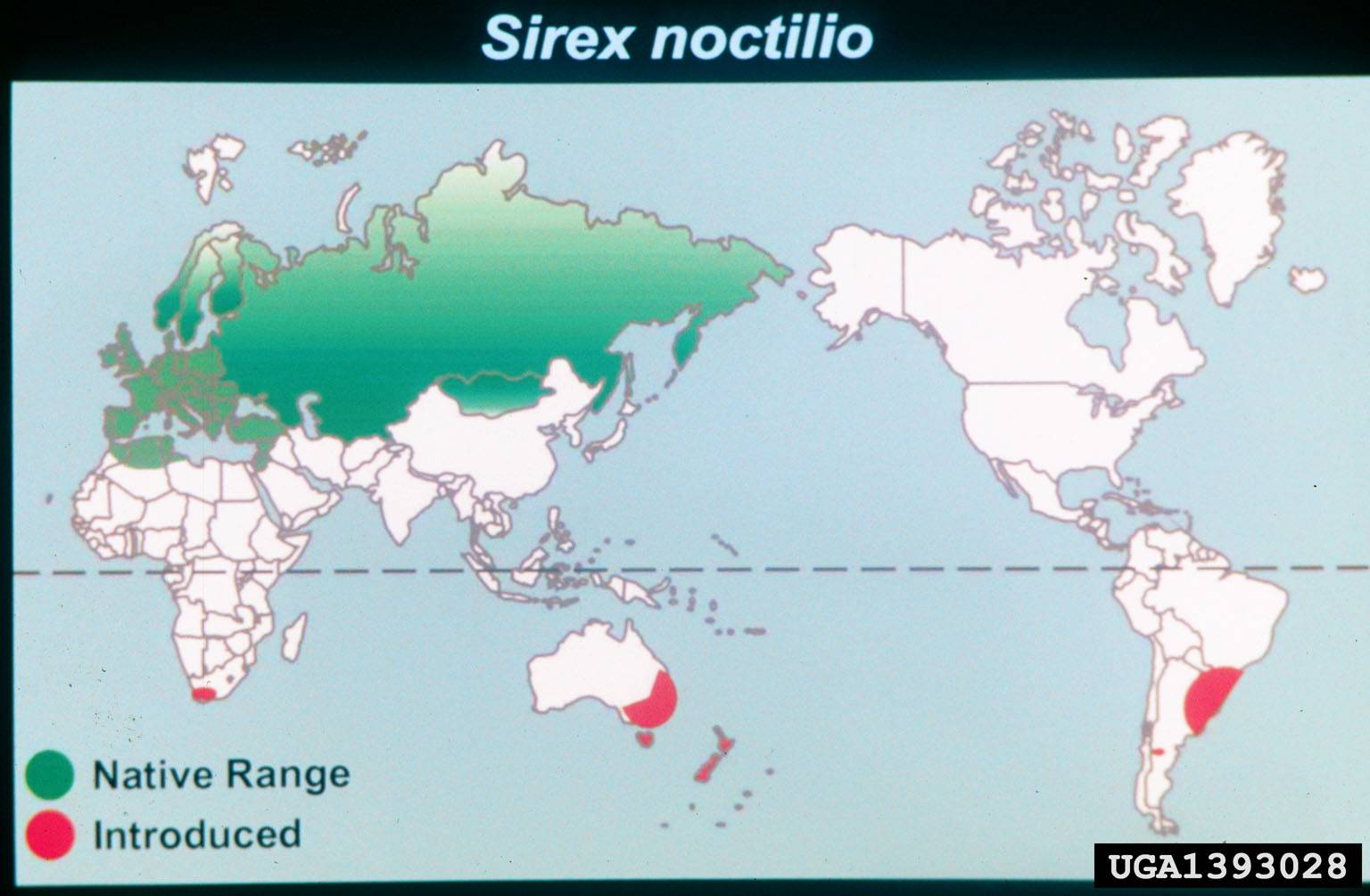









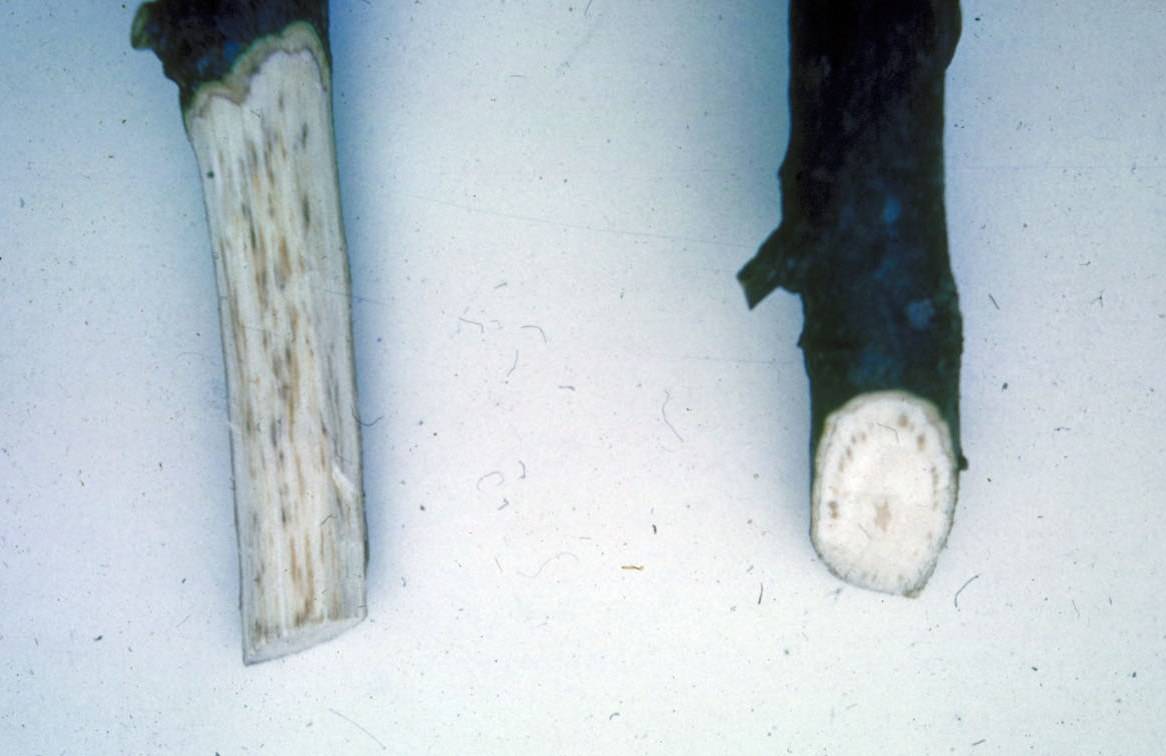









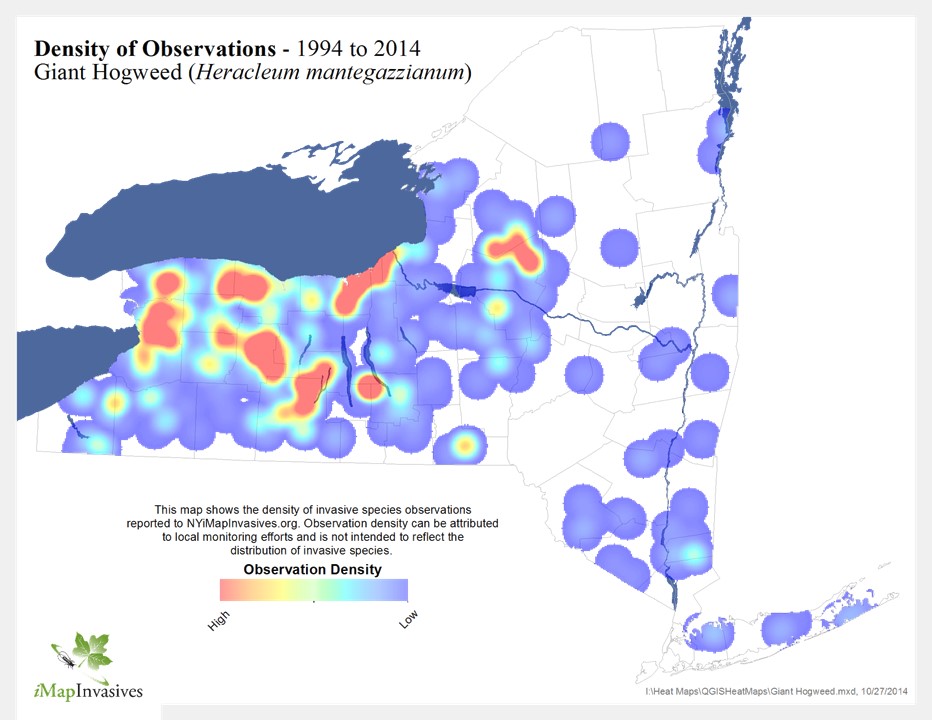




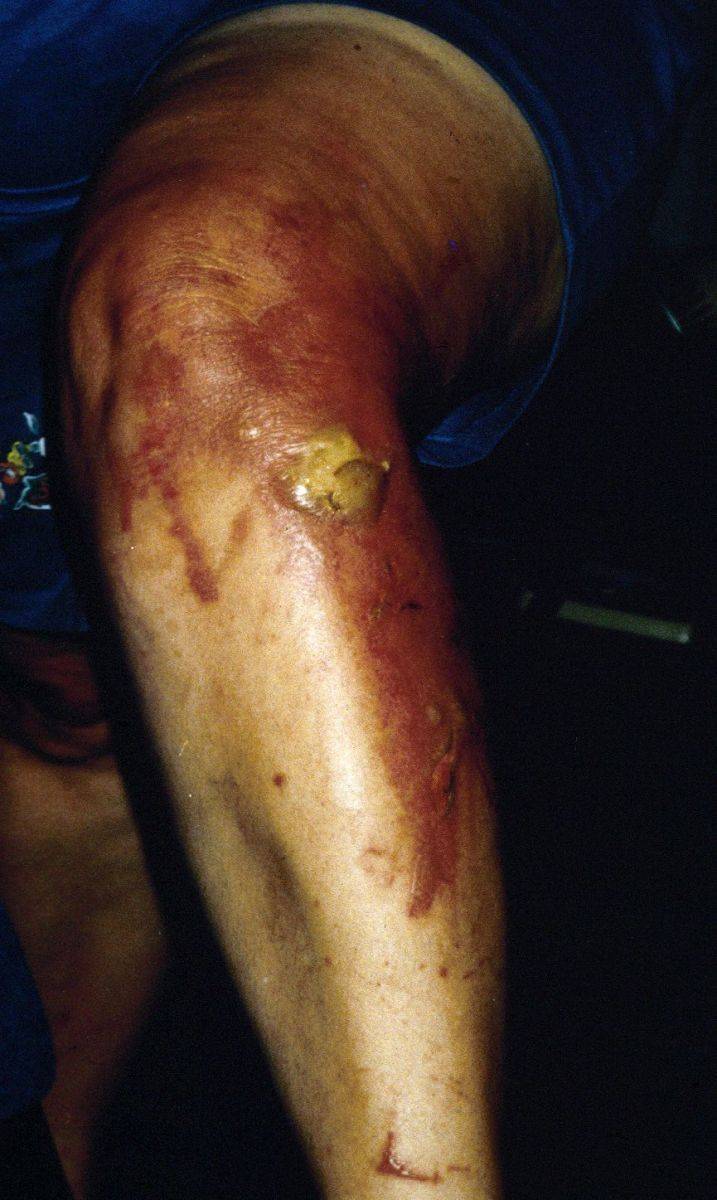













.jpg)


