The spring ephemeral lesser celandine (Ficaria verna, previously Ranunculus ficaria L.), also called fig buttercup, is an herbaceous perennial found throughout the northeastern United States. It prefers moist, sandy soil, and thrives along stream and river banks, in open forested flood plains, and in other wetland sites. It will grow in drier habitats, however, and as a garden escape can often be found in urban and semi-urban areas.
Lesser Celandine, a.k.a. Fig buttercup (Photo: David Nicholls, NatureSpot.org National Biodiversity Network Trust [NBN Trust], UK)
Origin and Distribution
Native to Europe, Asia and northern Africa, lesser celandine was likely introduced as an ornamental plant. The earliest herbarium specimen dates to 1867 from Pennsylvania. Lesser celandine has been reported throughout the northeastern United States and west to Missouri, and in the Pacific Northwest.
Lesser Celandine US distribution (Map: EDDMapS. 2015. Early Detection & Distribution Mapping System. The University of Georgia – Center for Invasive Species and Ecosystem Health. Available online at http://www.eddmaps.org/; accessed October 1, 2015.
Identification
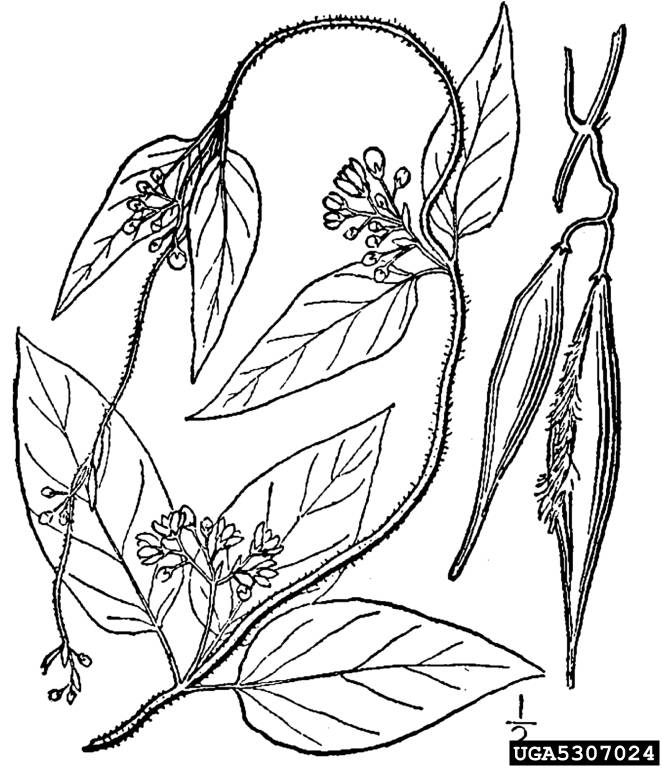
Lesser celandine is low growing and mat-forming, with long stalked leaved densely arranged in a basal rosette. Mature rosettes can reach up to 30 cm (12 in) diameter and up to 30 cm (12 in) tall. The kidney shaped leaves are 4 to 9 cm (1.6 to 3.5 in) wide, smooth, and have wavy edges. Leaf stalks (petioles) are deeply grooved and U-shaped in cross section. Flowers are yellow, 2 to 6 cm (0.8 to 2.4 in) wide and have long (10 to 30 cm; 4 to 12 in) peduncles. There are five sub-species of lesser celandine, distinguished largely by reproductive characters. Ornamental varieties may have a range of flower colors.
Close-up of flower petals and stamens – David Nicholls, NatureSpot.org National Biodiversity Network Trust [NBN Trust], UKBulbils formed at end of mature petioles allow for self-propagation – Antonino Messina, Flora of North AmericaTubrous roots – Andrea Moro, Dipartimento di Scienze della Vita, Università di Trieste
Biology
Lesser celandine is ephemeral (short-lived), and its emergence is triggered by increased light availability in the early spring. Shoots emerge from late-March to mid-April depending on environmental conditions, and flowering, which may be linked to water availability, occurs from late April to mid-May. Some sub-species produce pale aerial bulbils in the leaf axils. Seed production occurs in late spring, and by summer the above ground vegetation dies back and the plant becomes dormant. Seeds do not mature on the plant; rather, they require an after-ripening period to fully mature. Some varieties do not produce many viable seeds, and the aerial bulbils and tuberous roots are the primary mode of reproduction. Bulbils are dispersed through animal and environmental disturbance, and can be carried along waterways.
Lesser celandine plant structure (Illustration: Duna-Ipoly Nemzeti Park Igazgatosag, Hungary, at Dinpi.hu)
Impacts
Lesser celandine emerges earlier than most native plants, and may inhibit the development and reproduction of other spring ephemerals, particularly smaller species like spring beauties (Claytonia spp.), trilliums (Trillium spp.), and bloodroot (Sanguinaria canadensis). These native wildflowers are a source of nectar for bees and other insects in the early spring. The bare ground left behind after lesser celandine senesces in late spring may be colonized by other weedy species.
Lesser celandine taking over a forest understory. (Photo: Les Merhoff, DiscoverLife.org)
Control and Management
Due to its ephemeral nature and capacity for vegetative reproduction, lesser celandine can be challenging to effectively control. Small infestations can be removed manually, though care must be taken to completely remove the tubers from the soil. Repeated, early season mowing may reduce or stay growth of lesser celandine, but risks dispersing aerial bulbils. Glyphosate based herbicides may also be effective, and will require far less soil disturbance than manual removal. A 1.5% concentration of a 39% to 41% glyphosate isopropyl-amine salt solution with a non-ionic surfactant is effective for spot applications. Herbicide is most effective when applied early in the season and, when possible, applications should be made before any non-target plants have emerged. Note: Always check state/provincial and local regulations for the most up-to-date information regarding permits for control methods. Follow all label instructions. Mention of chemicals in this profile does not represent a recommendation by NY Sea Grant or Cornell University.
New York Distribution Map
This map shows confirmed observations (green points) submitted to the NYS Invasive Species Database. Absence of data does not necessarily mean absence of the species at that site, but that it has not been reported there. For more information, please visit iMapInvasives.
Wineberry (Rubus phoenicolasius) is an invasive shrub in the same genus as raspberries and blackberries. Wineberry creates spiny, inpenetrable thickets that reduce an area’s value for wildlife habitat and recreation. It was introduced to North America in the 1890s as breeding stock for raspberries. It was found invading natural areas by the 1970s, and it is currently recorded in most states east of the Mississippi River and in Alabama (USDA PLANTS Database). Wineberry replaces native vegetation, including native edible berry shrubs. It is differentiated from other berry-producing canes by the reddish appearance of its stems (caused by a dense coat of red hairs), silvery underleaf surfaces, and bright red berries. Management can be obtained through mechanical, chemical, or combination of control methods.
Wineberry flowers. (Photo: Leslie J. Mehrhoff, University of Connecticut, Bugwood.org)
Wineberry flowers. (Photo: Leslie J. Mehrhoff, University of Connecticut, Bugwood.org)
U.S. range of wineberry. (Map: USDA NRCS PlantsDatabase)
Biology and Habitat
Wineberry is a close relative of other raspberries and blackberries. It grows in long shoots called canes up to six feet long, which can re-root at the tips when they touch the ground. Wineberry canes grow in two stages; in the first year they form a vegetative cane, and in the second year the cane becomes woody and produces lateral branches, flowers, and fruit (technically drupes, an aggregation of single seeded drupelets, but for clarity the term fruit will be used). Wineberries are perennial; while the canes each live two years, the plant produces new canes every year. Leaves are produced in April, flowers in May, and fruit from late June to August; leaves drop in late November. Wineberry does not need pollen from another individual to set seed, and therefore may reproduce more easily than natives like saw-toothed blackberry (Foss 2005).
Wineberry has a wide range of tolerance for light, soil type, and moisture level, and is hardy to USDA Zone 5 (annual minimum temperatures to -20F). While it is most productive in edge and wasteland habitats, it can be found in most habitats that exist in New York (Innes 2009), including forested habitats. Wineberry seeds are spread by animals, and seeds dropped on the forest floor can germinate when falling trees provide light to the forest floor. Once established, wineberry can persist indefinitely and reproduce once further disturbance occurs (Innis, 2005).
Wineberry canes. (Photo: Leslie J. Mehrhoff, University of Connecticut, Bugwood.org)
Identification
Wineberry is related to other raspberries and blackberries, and shares characteristics of both. Like raspberries, wineberry has silvery underleaves, a fruit core that remains on the stem when the ripe fruit is picked, and thorns. It is differentiated from other raspberry species by the fine red hairs that grow densely on its stems (and flowers) causing a reddish hue to the plant. Wineberry fruit is vibrantly red when ripe, which helps differentiate it from native black raspberries and blackberries; it also has three leaflets per leaf rather than five, which separates it from many blackberry species. Unique to wineberry is its small, greenish, hairy flowers with white petals and the way its fruit remain covered by sepals (greenish petal-like structures) until almost ripe.
Wineberry stem hairs. (Photo: Leslie J. Mehrhoff, University of Connecticut, Bugwood.org)Top and bottom of wineberry leaves. (Photo: Leslie J. Mehrhoff, University of Connecticut. Bugwood.org)Wineberry hairs. (Photo: Ansel Oomen, Bugwood.org)
Rubus odoratus (purple-flowering raspberry or thimbleberry) has maple-shaped leaves that are soft and hairy; leaves not silvery; flowers pinkish-purple. Fruit is flatter and fuzzier than a raspberry, forming more of a cup shape.
Black raspberry (Rubus occidentalis) has whitish underleaves, but flowers hold their white petals out from the center of the flower, and fruit are usually purple-black (occasionally golden). Stems are green with a bluish cast that rubs off and have sparse, fairly robust thorns. Canes tip-root.
Red raspberries (Rubus idaeus, Rubus strigosus and many hybrids) have whitish underleaves and white petals, with red fruit, like wineberries. Stems are not covered in red hairs, are more lightly armed than black raspberry, and lack the bluish-white cast on their stems. Flowers might have a few hairs, but are not densely hairy like those of wineberry.
New York has several species of native blackberries, all of which have green rather than silvery underleaves and solid-cored fruit (mostly black when ripe). Some have five to seven leaflets. Identification to species can be difficult. While the skin on some species is reddish or purplish, none are covered in reddish hairs like wineberry, and many are heavily armored with thorns.
Black raspberry (Rubus occidentalis) leaves and canes. (Photo: D. Cameron, from Go Botany website: https://gobotony.newenglandwild.org)
Evergreen blackberry (Rubus laciniatus) is an invasive blackberry. It has highly dissected leaves and black fruit with a solid core.
Evergreen blackberry canes and leaves. (Photo: Joseph M. DiTomaso, UC Davis. Bugwood.org)Evergreen blackberry leaves and unripened fruit. (Photo by Joseph M. DiTomaso, University of California – Davis, Bugwood.org)
Himalayan blackberry (Rubus armeniacus) is also an invasive blackberry. It has stout, heavily armed but not hairy stems that grow up to 20 feet, tip roots like wineberry does, and produced large, sweet, dark-purple to black solid-cored fruit. It is the only blackberry with a whitish or grey-green underleaf, but usually has five leaflets instead of three, which along with its pinkish-white flowers and black fruit differentiate it from wineberry.
Himalayan Blackberry canes. (Photo: Joeph M. DiTomaso, UC Davis. Bugwood.org)
Ecological Impacts
Wineberry can form dense, impenetrable thickets in natural areas, making the habitat unusable for some species and creating hiding places for others. It is more aggressive than many of the native raspberry and blackberry species, and has a wider range of tolerance for light, soil type, and moisture. Its establishment in forest understories as disturbance occurs can lead to its spread even in mature forests. There has been no study to date documenting its specific impact on native species.
Wineberry plants choking understory of second growth forest. (Photo: John M. Randall, The Nature Conservancy. Bugwood.org)
Control
Wineberry control is more straightforward than control of many other invasive plants in New York. While any root fragments may start a new plant, wineberry does not have a vigorous underground storage structure; this makes it easier to control than, for instance, Japanese knotweed or lesser celandine. It is also susceptible to common pesticides.
For any invasive species control project, it is important to have a plan for the location before control begins. Disturbance without replanting often results in the return of either the same invasive species or other invasives to the site; have a restoration plan in place before starting invasive species removal.
Mechanical control
Hand pulling wineberry or digging with a spading fork can be a successful strategy in small patches or where repeat visits are not costly, particularly if native species are planted where the ground has been disturbed. Return visits for a few years will be necessary to remove new plants that sprout from root fragments. As wineberry is armed with thorns and hairs, minimizing exposed skin during mechanical control is advisable.
Chemical control
Wineberry can be controlled using systemic herbicides such as glyphosate or triclopyr (Bargeron et. al., 2003). When using pesticides, be aware that many pesticides are prohibited within 100’ of water, as they are toxic to aquatic life and/or fail to break down in water. Some formulations of glyphosate-based herbicides are permitted for use near water, but the most common formulation (Roundup) is not permitted for use near water due to an adjuvant (chemical that helps the glyphosate stick to plant surfaces) that is toxic in aquatic habitats. Triclopyr also has both aquatic-permitted and prohibited formulations; choose carefully based on the characteristics of your treatment area. Always follow instructions on the label of any pesticide, and remember that New York has its own regulations for pesticides, both for the entire state and for specific regions like Long Island that have special environmental considerations. For New York State regulations, visit the DEC website: http://www.dec.ny.gov/regulations/8527.html.
Foliar application and cut-stump application are both recommended in various fact sheets (Massachusetts Audubon, Innes 2009, bugwoodwiki), but no experiments have been published on the relative efficacy of pesticides or application methods on wineberry (2015).
New York Distribution Map
This map shows confirmed observations (green points) submitted to the NYS Invasive Species Database. Absence of data does not necessarily mean absence of the species at that site, but that it has not been reported there. For more information, please visit iMapInvasives.
Japanese virgin’s bower (Clematis terniflora DC, also called sweet autumn clematis) is an invasive ornamental vine in the Clematis genus. It is widely planted in gardens for its profusion of small, showy white flowers in late summer and equally showy silvery, corkscrew, feathery seedheads. Given support it can reach 30’ in height. Its prolific nature and aggressive growth make it a problem in natural areas, where it can smother mature trees and displace native vegetation. Japanese virgin’s bower is on New York State’s regulated species list; it can be transported, purchased, sold and planted, but it can only be planted where it will not threaten natural areas, and must be displayed for sale with signs explaining that it is an invasive species.
Identification
Japanese virgin’s bower is native to Japan and China. It was brought to North America as an ornamental species. Leaves are opposite and compound, with three to five heart-shaped leaflets 2-3½” long; the surfaces are dark green and shiny, paler on the undersides. It climbs using tendril-like petioles to cling to support. Flowers are about 1” across, white, four-petaled, and held in clusters of 3-15; the plant blooms in late summer to fall. Seeds are small and brown, with showy, silvery-grey, feathery, curving seedheads.
Look-alikes
Japanese virgin’s bower looks like many other Clematis genus species; the closest match is a native plant also commonly called virgin’s bower, Clematisvirginiana. Flowers and seeds are similar between the two species, but leaves of C. virginiana are toothed and lobed as opposed to C. terniflora’s entire, ovate leaves. The other Clematis have larger flowers of various colors; these ornamental species and hybrids are not particularly aggressive.
Biology & Habitat
Japanese virgin’s bower can survive a wide range of soil types and light availability, although it is most productive with shade on its roots and full sun on its foliage. It establishes in hedgerows, waste spaces, roadsides, stream banks and forest edges. It is a semi-woody perennial, producing foliage on new stems each year. In southern climates it is semi-evergreen. Japanese virgin’s bower produces shoots from last year’s woody stems in spring. It flowers in August and produces seeds by late September, which are dispersed by wind.
Prevention & Control
Cultural: The similar native virgin’s bower, Clematis virginiana, has many of the same ornamental characteristics as Japanese virgin’s bower and is less aggressive. They are so similar they are sometimes sold interchangeably at nurseries. Planting a native or noninvasive alternative will reduce the chances of this invasive damaging nearby natural areas.
Monitoring: Japanese virgin’s bower is most easily seen in the late summer, when it produces a froth of white flowers. Otherwise, look for a vine with small, oval- to heart-shaped, opposite leaves. There is currently no research on control of this species; all information is from gardening and extension sources.
Manual and Mechanical Control: Remove the plants and its roots. Some root fragments may resprout, so repeat visits may be necessary. Given the plant’s prolific nature, expect seeds still in the soil to continue to sprout new plants for some time. Research on the longevity of C. terniflora’s seeds in the soil is not available.
Biological Controls: Currently there are no biological controls for Japanese virgin’s bower.
Chemical Controls: Japanese virgin’s bower responds to control with glyphosate and triclopyr and other common broadleaf herbicides, with no known resistance issues. However, all will require repeat treatments (Langeland and Meisenburg, 2009). Before using chemical controls, always check with your state’s environmental regulatory agency to check on the legality of using any particular chemical.
New York Distribution Map
This map shows confirmed observations (green points) submitted to the NYS Invasive Species Database. Absence of data does not necessarily mean absence of the species at that site, but that it has not been reported there. For more information, please visit iMapInvasives.
Kudzu, “the vine that ate the South.” Kerry Britton, USDA Forest Service, Bugwood.org
Introduction & Distribution
Kudzu (Pueraria montana) is a semi-woody, trailing or climbing, perennial invasive vine native to China, Japan, and the Indian subcontinent. Kudzu is also known as foot-a-night vine, Japanese arrowroot, Ko-hemp, and “the vine that ate the South.” The vine, a legume, is a member of the bean family. It was first introduced to North America in 1876 in the Japanese pavilion at the Philadelphia Centennial Exposition. A second major promotion of kudzu came in 1884 in the Japanese pavilion at the New Orleans Exposition. The first recorded use of kudzu in North America was as a shade plant on porches in the American South (the plant produces attractive, fragrant purplish flowers in mid-summer). Kudzu was heavily promoted in the early-1900s when the government paid farmers to use the vine for erosion control (more than a million acres are estimated to have been planted as a result) and as a drought-tolerant, nitrogen-fixing legume (capable of bacterial growth with stem and root nodules converting free nitrogen to nitrates, which the host plant utilizes for its growth in low nitrogen soils) for livestock feed. During the Great Depression, thousands of acres of kudzu were planted by the Civilian Conservation Corps for hillside stabilization projects. In some areas, kudzu blossoms have been prized for their use in making kudzu blossom jelly and jam. The long kudzu fibers are also used in basket making. Ko-hemp, a more refined version of kudzu fiber has long been used for cloth weaving in China.
Use of kudzu for cattle grazing in the early-1900s. USDA NRCS Archive, USDA NRCS, Bugwood.org
These government-sanctioned uses of the vine, combined with its innate, aggressive, range-expansion capabilities resulted in a rapid spread of kudzu throughout North America. Kudzu can now be found in 30 states from Oregon and Washington State to Massachusetts, particularly infesting states from Nebraska and Texas eastward most heavily; the vine is most common in the South. It has also been discovered in Hawaii and the warm, south-facing growing region on the north shore of Lake Erie in the Canadian Province of Ontario.
U.S. range of kudzu. USDA PLANTS database, July 2014.
Biology & Identification
Kudzu is an herbaceous to semi-woody, climbing or trailing, nonnative, deciduous, perennial vine or liana (a vine that is rooted in ground-level soil and uses trees and other vertical supports (telephone polls, buildings, etc.) to climb to the forest canopy to get access to light. A well-known example would be common wild grape).
Kudzu covering other vegetation and forming a liana. James H. Miller, USDA Forest Service, Bugwood.orgClose-up of a kudzu liana. Leslie J. Mehrhoff, University of Connecticut, Bugwood.org
Kudzu produces long, hairy vines from a central root crown. Kudzu has dark-green, hairy, alternate, compound leaves, 2 – 8 inches (5 – 20 cm) in length with three oval- to heart-shaped leaflets 3 – 4 inches (8 – 10 cm) long at the end; these leaves may be slightly or entirely lobed. Stems are also hairy. Vines can grow up to 30 to 100 feet (9 – 30.5 meters) per year. The vines have 0.8 – 1 inch (2 – 2.5 cm) flowers on 4 – 8-inch (10 – 20 cm) axillary racemes (short, equal length stalks along a main stem forming clusters of flowers with the oldest flowers toward the base with the newest end of the stalk terminating in one or more undeveloped buds). Vertical kudzu vines in full sunlight produce flowers in late-summer; horizontal vines seldom produce flowers. The flowers are typically red, purple, or magenta with a strong, grape-like aroma; pink or white flowers occur occasionally.
Kudzu leaflets found at end of stem. James H. Miller, USDA Forest Service, Bugwood.orgKudzu flowers made the plant popular for planting around porches. Forest and Kim Starr, Starr Environmental, Bugwood.org
Population Expansion
Kudzu populations spread both asexually and by seed germination.
Asexual (vegetative) spread:
The most common method of spread is by setting new root crowns at almost every node where horizontal trailing stems come in contact with bare soil (this can be every few feet); new vines will form at these nodes the following spring and will spread out in all available directions. Kudzu tap roots can grow up to 12 feet (3.6 meters) long and weigh up to several hundred pounds. This may help kudzu to withstand long periods of drought.
A typical mature kudzu root crown. The Coalition To Control Kudzu Without Chemicals (http://www.kokudzu.com/)
Sexual spread:
Kudzu usually does not flower until its third year, with flowers and seeds forming only on vertical climbing vines. Kudzu produces clusters of 20 – 30 hairy brown seed pods, 1.6 – 2 inch (4 – 5 cm) long pods. Each pod contains from 3 to 10 kidney bean-shaped seeds, of which only 1 or 2 seeds are viable. Dormant viable seeds are unable to germinate until after their seed coats have become water permeable as a result of physical scarification (breaking the seed coat by abrasion or prolonged thermal stress). Seeds deposited below the vines in the seed bank may take several years to germinate. This can be problematic during control efforts because it can result in the reemergence of the plants years after eradication was believed to have been achieved. It has been observed that kudzu in North America is more likely to grow asexually than by setting seed. It appears that this is due to kudzu seedlings being outcompeted by vegetatively produced vines.
A small cluster of kudzu pods. James H. Miller & Ted Bodner, Southern Weed Science Society, Bugwood.org
Habitat & Ecology
Factors that help determine how invasive kudzu will be in any habitat appear to be climate and availability of light. Warmth and humidity are important factors, with greater colonization corresponding to warmer average annual temperatures and higher average humidity. To reach additional light, the vines climb existing vegetation and hard vertical surfaces. It does not appear that the composition of the local native plant community has much influence on kudzu invasiveness. Even undisturbed plant communities adjacent to an existing population of kudzu can be at risk. Typical kudzu habitats are usually open, disturbed areas such as roadside ditches, rights-of-way, and abandoned fields. In such settings, kudzu can form large monocultures with thousands of plants per acre.
Kudzu has a strong daily leaf orientation capability; by controlling the leaf position as it faces toward or away from the sun, kudzu can control sunlight intensity on the leaflets that are exposed. This ability can reduce leaf temperatures relative to native vegetation and minimize the amount of water lost from the plant by leaf surface transpiration during times of peak sunlight. It may also be a benefit below forest canopies where light is dim by increasing the surface area of leaves receiving sunlight. Leaves exposed to open sunlight may be able to maximize photosynthesis, store additional food in kudzu’s rhizomes, and have a competitive advantage over native vegetation.
Kudzu accumulates and maintains substantial carbon reserves in large woody, tuberous roots, again giving it a competitive advantage.
Trailing stems in open areas tend to die back in the winter. Vertically climbing vines develop thick bark and can reach diameters greater than 0.8 inch (2 cm), aiding in overwintering.
Kudzu vines can more easily grow around smaller vines such as honeysuckle (Lonicera spp.) than around bare tree trunks. This growth tactic appears to aid the plant in the formation of lianas in forested areas. Once established, kudzu lianas compete with forest trees both for sunlight in the crown and for water and nutrients from the soil. The vines may directly damage colonized trees by strangulation. These physical traits of a kudzu liana significantly impact the ability of native trees to grow and reproduce, increasing the early mortality of native trees, and preventing the establishment of new trees or shrubs in the dim light below the colonized canopy.
Kudzu lianas can cause weakened trees to fall from the weight of the overgrowth of vines or by pulling down trees attached to the liana when one weak tree succumbs to the weight of ice freezing onto the tree and/or the vines.
Kudzu thrives where the climate favors mild winters (40 – 60°F {4 -16°C}), summer temperatures rising above 80°F (27°C), precipitation greater than 40 inches (101 cm), and a long growing season.
Because of its underground root crowns, kudzu can escape fire damage. During the growing season, kudzu’s underground root system can provide significant water to the foliage; the high water content stems and foliage are able to resist some fire damage that may kill nearby native plants.
There is some indication (not yet definitively proven) that wildfire (or controlled burn) soil heating may promote kudzu seed germination by scarifying the seedcoat which would allow penetration by water to allow for germination.
Robert L. Anderson, USDA Forest Service, Bugwood.org
Impacts
Native Plant Community Impacts:
A kudzu invasion can cause several different types of major impacts on native plant communities: it can crowd them out; it can outcompete them; and it can physically crush them.
Since kudzu can fix nitrogen in its roots, it can thrive in soils too low in nitrogen to support robust growth of native vegetation, thereby outcompeting native plants for both nutrition and growing space, ultimately forming monospecific plant communities. This significantly alters natural plant communities and the animals that rely on those natural communities for food and habitat. Areas of more than 100 acres (40 hectares) with 1 – 2 plants per square foot, or 40,000 to 85,000 plants per acre (107,000 to 215,000 plants per hectare) can be found in the American South.
Kudzu’s rapid growth rate and its manner of growing over whatever it encounters in its path can also overwhelm native plant communities, also resulting in monospecific stands of the vine.
As heavy infestations of kudzu can completely cover trees of almost any size, kudzu lianas can both fell trees from their extreme weight or nearly eliminate light availability within the forest canopy, weakening or killing shade-intolerant species, particularly pines. Once kudzu gains access to the forest canopy, the liana formed can spread faster and more aggressively through a forest.
An extreme example of kudzu overgrowth of natural vegetation. Kerry Britton, USDA Forest Service, Bugwood.org
Economic Impacts:
By outcompeting, smothering, and physically removing native vegetation, kudzu damages to lost forest production for southern commercial timber producers has been estimated to be as high as $48 per acre ($118 per hectare) per year. Kudzu control costs can be as high as $200 per acre per year. Control costs on power company rights-of-way and transmission equipment have been estimated as high as $1.5 million per year. Kudzu can also be a problem along highway rights-of-way.
Kudzu overgrowth of a southern highway embankment. Chris Evans, Illinois Wildlife Action Plan, Bugwood.org
Control
With a growth rate of up to one foot (0.3 meter) per day, simply controlling or managing kudzu can become a “fool’s errand” of never ending activity. In areas where the plant cannot be tolerated at all, kudzu control is basically kudzu eradication. To prevent reinvasion, complete eradication is required, which means every root crown on a site must be killed. Due to the numerous root crowns at vine nodes, eradication of a well-established population of kudzu could take 5 – 10 years of concentrated effort. The more mature the population, the more difficult eradication becomes as a result the numerous crowns and the large rhizome system that can store significant amounts of starch to feed the plant. Lianas are also more efficient at producing starch and sending it to the root system than are horizontal, ground-based vines.
Eradication of kudzu with herbicides calls for frequent defoliation during the growing season, while most of the plant’s energy is devoted to vine production and growth. Defoliation forces the plant to call on root starch reserves to resume foliage growth activities, helping to diminish reserves of starch and prevent storage of new reserves. If a single treatment is all that can be undertaken in a year, it should be implemented in early-fall as foliage starch allocation to the root system replenishing that used for growth during the spring and summer takes place in the early-fall.
If physical or mechanical control methods are selected, eradication of well-established kudzu populations could take many years or be ineffective in the long-term. Mechanical harvesting of kudzu foliage limits the production of new food reserves by reducing photosynthesis; regrowth helps to deplete starch stored in the root system. Mowing of trailing vines and root crowns every two weeks may take up to ten years to eradicate small, immature patches of kudzu, assuming that all root heads are mowed. Mowing is more likely to result in eradication if used with herbicide application. During mechanical eradication efforts, all cut plant material should be destroyed by burning or by bagging and landfilling.
The use of intensive conservation grazing by herbivores such as sheep or goats can help control young, tender kudzu growth and make control by herbicides more effective over shorter periods of time by helping to reduce energy reserves.
For information regarding appropriate use of herbicides against kudzu and other invasive plants, please consult The Nature Conservancy’s Weed Control Methods Handbook. Make certain to consult your state’s environmental conservation or natural resource management agency to determine which herbicides are legal for kudzu control in your state.
Herbicide spraying for kudzu control. James H. Miller, USDA Forest Service, Bugwood.org
Policy Status
In the 1950s, the Agricultural Conservation Program removed kudzu from the list of species acceptable for use as an agricultural forage crop or soil stabilization plant. Congress listed kudzu as a Federal Noxious Weed in 1998. In 2014, the State of New York designated kudzu as a prohibited plant under the state’s Environmental Conservation Law.
New York Distribution Map
This map shows confirmed observations (green points) submitted to the NYS Invasive Species Database. Absence of data does not necessarily mean absence of the species at that site, but that it has not been reported there. For more information, please visit iMapInvasives.
Wild parsnip (Pastinaca sativa). Leslie J. Mehrhoff, University of Connecticut, Bugwood.org
Introduction & Distribution
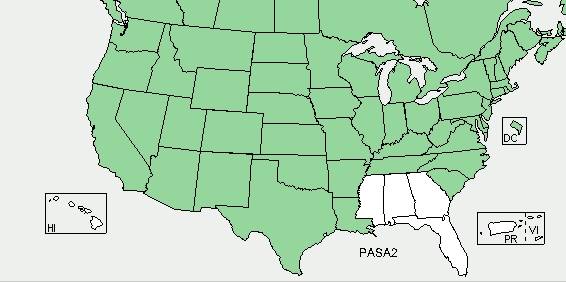
Wild parsnip (Pastinaca sativa) is a biennial/perennial herb native to Eurasia. In appearance, it looks and smells quite like cultivated parsnip (in point of fact, wild parsnip is part of the Apiaceae (or Umbelliferae) family which includes carrots, celery, parsley, parsnip, Angelica, and Queen Anne’s Lace, most of which are aromatic plants with hollow stems). It is believed to be an escapee from parsnip that was originally under cultivation. The plant typically can grow up to 4 feet (1.2 m) tall in an average year. Wild parsnip is common throughout the northern United States and southern Canada. Its range reaches from Vermont to California and south to Louisiana (it is not found in Hawaii, Mississippi, Alabama, Georgia, and Florida). Reported populations can be found across New York State with the heaviest concentrations being found in the Lower Hudson Valley, Catskills, and southern Adirondacks.
North American range of wild parsnip (Pastinaca sativa). USDA PLANTS database, July 2014.
Habitat, Biology & Ecology
Wild parsnip is an herbaceous plant which can grow from 4 – 5 feet (123 – 150 cm) tall. It can survive in a broad range of environmental settings, from dry soils to wet meadows. It grows best in rich, calcareous, alkaline, moist soils. It is commonly found growing along roadsides, in pastures, and in abandoned fields, or any place where the soil has been disturbed and native vegetation has yet to become fully established.
The roots are generally smooth and cylindrical, although sometimes lateral roots will grow out from the central tap root.
Seedlings emerge from February through April, form rosettes in their first year, and grow vegetatively for one or more years, at which time they will form an aerial shoot (called a “bolt”) and flower. Wild parsnip produces a rosette of broad, hairless, ovate, compound pinnate leaves, up to 6 inches (15.2 cm) in length, terminating with several pairs of leaflets with saw-toothed margins; they can grow up to 16 inches (40 cm) long. Leaflets are arranged in pairs along the stalk. Lower leaves have short stems, upper leaves are stemless. The leaves give off a pungent odor when crushed. During the vegetative growth season, wild parsnip continuously produces and loses leaves. The flower stalk develops from the rosette in the second year and can grow to a height of 4 – 5 feet (123 – 150 cm). It is grooved, hairy, and, except at the nodes, hollow. The stalk is sparsely branched. Over the winter above ground wild parsnip plants die back with only one or two leaves remaining on each plant.
First-year wild parsnip growth. Patrick J. Alexander @ USDA-NRCS PLANTS Database.
Close-up of wild parsnip leaves. Patrick J. Alexander @ USDA-NRCS PLANTS Database
Wild parsnip stems are hollow except at the nodes. Photo: Missouri Dept. of Conservation
Wild Parsnip stem. Leslie J. Mehrhoff, University of Connecticut, Bugwood.org
Each wild parsnip plant produces hundreds of small yellow flowers which bloom from June to mid-July. The flowers are arranged in a loose compound umbel (a structure made up of a number of short flower stalks which spread from a common point, looking like the ribs of an umbrella). An umbel can measure from 4 – 8 inches (10 – 20 cm) in diameter. The flowers consist of five yellow petals curled inward, five stamens, and one pistil. The large, straw to light-brown seeds that are produced by the flower heads are round to oval, flat, slightly ribbed with narrow wings and are 1 ½ – 3 inches (4 – 8 mm) long. Seeds mature by early July. Plants die after producing seeds; the dead stalk will remain standing through the winter. Seeds can remain viable in the soil for four years. Seedling mortality is high; less than 1% of seedlings survive to mature and reproduce.
Wild parsnip second-year growth. Photo: Virginia Tech Weed ID Guide
Mature flowering wild parsnip. Photo: University of Massachusetts Extension
Wild parsnip umbel. Virginia Tech Weed ID GuideWild parsnip seeds. Leslie J. Mehrhoff, University of Connecticut, Bugwood.org
Click the table (above) for a 2-page ID guide to giant hogweed, native cow parsnip, native purple-stemmed Angelica, poison hemlock, and wild parsnip
Impacts
Ecological Impacts:
Wild parsnip invades and modifies disturbed open habitats. Well-established fields and meadows are not likely to be invaded, but parsnip can become well-established along the edges and in disturbed areas. Once an infestation begins, it can spread into adjacent areas and form dense stands in high-quality fields and meadows. Wild parsnip is also very persistent on sites that remain disturbed or bare such as paths, roadsides, and utility rights-of-way.
Human Health Impacts:
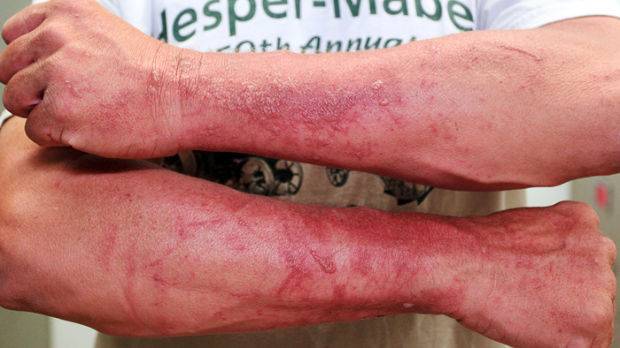
While wild Parsnip roots are edible, the plant produces a compound in its leaves, stems, flowers, and fruits that causes intense, localized burning, rash, severe blistering, and discoloration on contact with the skin on sunny days. This condition, known as phytophotodermatitis, is caused by furanocoumarin contained in the sap. This is not an allergic reaction, it is a chemical burn brought on by an increase in the skin’s sensitivity to sunlight. Affected areas can remain discolored and sensitive to sunlight for up to two years, similar to but not as severe as contact with giant hogweed. This reaction is not brought on by contact with the foliage of the plant, only by contact with the sap.
Contact may occur when working, hiking, and harvesting crops, including when visiting u-pick operations. To reduce the risk of exposure to wild parsnip sap, when undertaking such pursuits one should wear long-sleeved shirts, gloves and long pants.
If one should come in contact with wild parsnip sap, you should immediately cover the exposed skin to prevent the reaction to sunlight (but the area will remain sensitized for about eight hours). The contact area should be washed with warm water and a mild soap. If exposure to sunlight causes a burn and blisters to develop the affected area should be covered with a cool, damp cloth to help relieve pain. The blistered skin should be kept out of the sunlight to avoid further burning. If blistering is severe, see a physician. There is no cure for parsnip burns; however, a topical or systemic cortisone steroid may relieve discomfort.
Wild parsnip burns. Andrew Link, Lacrosse Tribune 2013
The essential oil of parsnip roots contains a large percentage of Myristicine, a strong human hallucinogen.
Wild parsnip is regulated in Ohio, Illinois, Tennessee, and Wisconsin.
Prevention of Establishment & Spread
Remove new infestations while they are still small. Avoid mowing areas with wild parsnip when viable seeds are present as equipment readily spreads seed to new areas. Clean mowing equipment before moving from an area with wild parsnip to one without. When possible, plan to harvest/mow areas without wild parsnip before moving to fields where it is present. Time control efforts to prevent spread of the plant.
Control & Management
Management decisions should be based on the quality of the area, the degree of the infestation, and use of the infested area by people or livestock.
Manual control for small patches is effective. Cut the root 1” below the ground using a tool such as a spaded shovel or remove plants by hand pulling, gripping the stalk just above the ground. These control measures should be undertaken before wild parsnip plants go to seed. If hand pulling after seed formation, take steps to destroy the seeds. For small areas which have set seed, cut the tops with clippers, bag the seed heads in clear plastic and allow to rot.
Mowing – Mow when plants first produce flowers, but before seeds enlarge. At this stage plants have depleted their root resources and often die when cut. Some plants will re-sprout, so a follow-up mowing may be needed. When using any type of mowing equipment, take precautions to prevent plant sap from contacting exposed skin. Mowing can tend to favor wild parsnip rosettes as more sunlight is able to reach them, as well as reducing the number of plants competing with them for light and nutrients.
Chemicals – General-use herbicides such as glyphosate or triclopyr can be applied as spot treatments to basal rosettes. Be sure to follow all label and state requirements.
Biocontrol – No effective options are currently known. The parsnip webworm infests individual plants, but is not known to significantly damage large patches.
Plan to monitor the area long-term for seedlings emerging from the seed bank.
Whatever type of control method is employed, make certain to take measures to protect skin and eyes from contact with the plant’s sap.
New York Distribution Map
This map shows confirmed observations (green points) submitted to the NYS Invasive Species Database. Absence of data does not necessarily mean absence of the species at that site, but that it has not been reported there. For more information, please visit iMapInvasives.
Mugwort (Artemesia vulgaris) is an invasive perennial forb that is widespread throughout North America, though it is most common in the eastern United States and Canada. It is a weed of nurseries, turfgrass, vineyards, waste areas, forest edges, and roadsides. Mugwort spreads aggressively through an extensive rhizome system and will readily form large, mono-specific stands.
Origin and Expansion
Mugwort is native to Europe and eastern Asia, where it has historically been used as a medicinal herb. Seed may have been first introduced to North America as early as the 16th century by Jesuit missionaries in Canada. It was also introduced throughout the continent as a contaminant in ship ballast and nursery stock.
Biology
Mugwort is a perennial with an extensive rhizome system. Shoots emerge during the spring, and flowering occurs from July to late September. A single plant can, depending on its environment, produce up to 200,000 seeds. The small seeds (~1mm in diameter) are largely wind dispersed. Seed production does not seem to be a major factor in the spread of mugwort populations, however, and some biotypes do not produce viable seed. Instead, mugwort spreads largely through vegetative expansion and the anthropogenic dispersal of root propagules. The root system is extensive though shallow (to 20 cm in depth), with numerous branching rhizomes up to 1 cm in diameter. Plants can regenerate from rhizome fragments as small as 2 cm (Klingeman et al. 2004).
Characteristics and Identification
The rarely-seen seedlings have oblong cotyledons without petioles. Adult stems are smooth and longitudinally ridged, with numerous axillary branches towards the upper portions of the plant. The stems become somewhat woody as they age. The leaves are alternate, densely covered with wooly, silver-white hairs on the underside, and slightly hairy on the upper surface. Leaf morphology is variable throughout the plant. The lower leaves are petiolate, with stipules at the base, and generally coarsely toothed and pinnately lobed. The upper leaves are sessile and lanceolate with smooth or toothed margins. The numerous ray and disk flowers are small (5 mm), green, and grow in racemes and clusters at the end of stems and branches. The foliage is aromatic and slightly pungent.
Impacts
Mugwort is a problematic weed in nurseries, where small root fragments can easily contaminate nursery stock. It is also a major weed in turf grass, field-grown ornamental crops, and orchards. Stands of mugwort displace native species, and can delay or disrupt succession in natural ecosystems (Barney and DiTommaso 2003). Mugwort produces several terpenoid potential allellochemicals, and decaying mugwort foliage has been shown to inhibit the growth of red clover in laboratory experiments (Inderjit and Foy 1999). Mugwort pollen is a common cause of hay fever.
Prevention, Control, and Management
The dense root system of mugwort can make it difficult to control. Pulling is ineffective, and may even promote growth by leaving residual rhizome fragments in the soil. Mugwort tolerates mowing, and even sustained mowing over two years will not fully eradicate mugwort stands. The relatively shallow roots make mugwort vulnerable to repeated cultivation in agricultural systems, though this practice risks spreading root propagules.
Chemical control of mugwort can have limited effectiveness. Though non-specific broadleaf herbicides such as glyphosate or dicamba can effectively control mugwort, the rates required for adequate suppression are rarely economical (Bradley and Hagood 2002). For small infestations, multiple spot-treatments of glyphosate can be effective (Bing 1983).
New York Distribution Map
This map shows confirmed observations (green points) submitted to the NYS Invasive Species Database. Absence of data does not necessarily mean absence of the species at that site, but that it has not been reported there. For more information, please visit iMapInvasives.
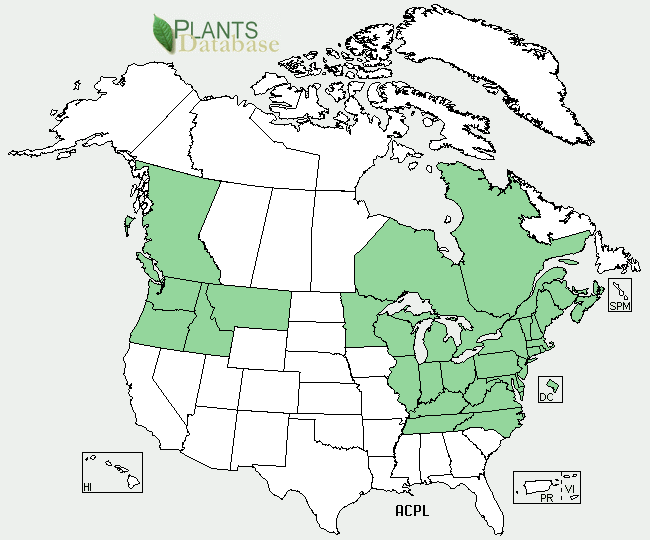
Norway maple (Acer platanoides) is a large deciduous tree that can grow up to approximately 40-60 feet in height. They are tolerant of many different growing environments and have been a popular tree to plant on lawns and along streets because of their hardiness. Norway maples have very shallow roots and produce a great deal of shade which makes it difficult for grass and other plants to grow in the understory below. Additionally, they are prolific seed producers and are now invading forests and forest edges.
Norway maple infestation. Photo: Leslie J. Mehrhoff, University of Connecticut, Bugwood.orgAdult Norway maple tree. Photo: Leslie J. Mehrhoff, University of Connecticut, Bugwood.org
Origin
Norway maples are native to Europe and Western Asia.
Habitat
Norway maples are found growing all over the United States in many diverse environments. They are very well adapted to extreme soil types and found in hardy to USDA Zone 4. Norway Maples tolerate a wide range of temperatures. They do prefer full sun, but may also be seen in shady habitats. Also, this species is tolerant to sulfur dioxide pollution and ozone, so they can thrive in urban settings or forests. Norway Maples will be found in the United States anywhere from the border of Canada south to the Carolinas in early or late succession forests, wetlands, yards or gardens, or in disturbed open areas and roadsides.
Introduction and Spread
Many Norway maples made their way from Europe to the United States by being transplanted as ornamental specimens or by having individual seeds escape cultivation. These trees produce ample amounts of winged seed which are dispersed readily in the wind and germinate quickly. A suggested reason for the success of many invasive species is the absence of predators in the invaded habitat. A study by Cincotta et al. (2009) found that foliar insect and fungal damage was significantly higher on native sugar maples than on the invasive Norway maples. This could be one of the contributing factors to the Norway maple’s success.
Impacts
Forests that are intact are generally more likely to ward off invaders. However, Norway maple has been found to be very successful at establishing itself in a variety of conditions including mature, deeply shaded forests (Martin and Marks 2006). Due to the dense canopy of Norway maples, forest diversity is starting to decline because the excess shade they create inhibits the regeneration of sugar maples and other native seedlings. The shallow root system makes growing difficult for other native shrubs and wildflowers in the understory. In urban environments, the root systems also destroy pavement, requiring expensive repairs. Other species of flora and fauna, such as insects and birds, may indirectly be affected due to the change in resource diversity and availability. Norway maple is also susceptible to certain types of fungi, such as Verticillium wilt and anthracnose and may also serve as a host for aphids.
Identification
Norway maples have simple, green, and opposite leaves. Leaves of Norway maples usually are broader than they are high, about four to seven inches wide, with five prominent lobes. The bark of a Norway maple is grayish black and furrowed. This species tends to leaf out earlier in the spring than other maples and forms a broad-rounded crown. Yellow or greenish-yellow flowers are approximately 8 mm in diameter and are found in clusters that are present from April to May. During the summer, fruits mature into helicopter-like blades with wide-spreading wings. In the fall, leaves usually turn a pale yellow. One of the easiest ways to differentiate Norway maple from sugar maple is to cut the petiole (or leaf stalk) or vein and if a milky substances oozes out, it is a Norway maple. Also, bud tips of Norway maples are more blunt, whereas sugar maples are pointy and sharp.
The prominent five-lobed leaf of the Norway maple. Photo: John M. Randall, The Nature Conservancy, Bugwood.orgThe wide-spreading wings of the Norway maple fruit. Photo: Paul Wray, Iowa State University, Bugwood.orgFlower clusters of Norway maple. Photo: Robert Vidéki, Doronicum Kft., Bugwood.orgBroad and blunt bud of a Norway Maple. Photo: Rob Routledge, Sault College, Bugwood.org
Prevention and Control
It is recommended to plant alternative tree species that are native to this region to prevent further spread of Norway maple. Other maple species that are native to the Northeastern United States include red maple (Acer rubrum), sugar maple (Acer saccharum), and silver maple (Acer saccharinum).
Seedlings of Norway maple can be pulled from moist soil before they get too large. Other types of manual removal include digging out saplings and root systems or cutting down large trees. Girdling the trees by removing the bark layer (including the cambium) can also be performed, but is most effective in the spring. Leftover stumps can be ground out or new growth that develops from old stumps can be cut in future years. Some chemical herbicides, such as glyphosate or triclopyr may be useful for control, but contact your local extension office for more information on chemical application and always follow state requirements and the instructions on the label.
Occurrences
Norway maple is found on both the western and eastern sides of the United States. It ranges from Canada in the north and down south to the Carolinas.
Distribution of Norway Maple in the United States. States shaded green have Norway maple present. In each of the shaded states, Norway maple is considered invasive. Map: Plants.usda.gov
New York Distribution Map
This map shows confirmed observations (green points) submitted to the NYS Invasive Species Database. Absence of data does not necessarily mean absence of the species at that site, but that it has not been reported there. For more information, please visit iMapInvasives.
Pale swallow-wort or dog-strangling vine (Vincetoxicum rossicum; syn. Cynanchum rossicum) and black swallow-wort or black dog-strangling vine (Vincetoxicum nigrum; syn. Cynanchum louiseae) are herbaceous invasive plant species. They were originally placed in the milkweed family, Asclepiadaceae, but in recent years the Asclepiadaceae has been recombined with the periwinkle family, Apocynaceae. Pale and black swallow-wort are two distinct species, but do have a similar growth habit and many similar physical characteristics. Both species are perennial climbing vines that form extensive patches and overgrow and smother native vegetation.
Black swallow-wort intertwined on itself. Photo: Leslie J. Mehrhoff, University of Connecticut, Bugwood.org
a stand of black swallow-wort. Photo: Leslie J. Mehrhoff, University of Connecticut, Bugwood.org
A stand of pale swallow-wort. Photo: Leslie J. Mehrhoff, University of Connecticut, Bugwood.org
Pale swallow-wort intertwined on itself with seed pods present. Photo: Chris Evans, Illinois Wildlife Action Plan, Bugwood.org
Origin
Black swallow-wort is native to the western Mediterranean region. Pale swallow-wort is native to Ukraine and southeastern European Russia.
Habitat
Black and pale swallow-wort tolerate a variety of soils and environmental conditions. They are typically found in upland habitats and have been observed in rocky shores, agricultural and natural fields, and woodlands. Swallow-wort favors full sun, but may thrive in semi-shaded areas as well. Pale swallow-wort also can be found in densely shaded sites. These species are disturbance- and drought-tolerant and can quickly spread once established.
Introduction and Spread
Swallow-wort may have been originally brought to North America for horticultural purposes as an ornamental plant. The shoots of swallow-wort start to emerge from the soil in the spring. The plants flower in the early summer and produce seed pods towards late July and early August. A square meter stand of swallowwort is capable of producing 1000-2000 seeds per year, which are then dispersed by wind. Black and pale swallow-wort are capable of self-fertilization and are polyembryonic, meaning each seed may contain multiple seedling producing embryos (Lumer and Yost 1995; St. Denis and Cappuccino 2004). The main method of spread is by seed; however, if the main stem is damaged, the perennating buds on the root crown will activate to produce axillary tillers (Weston et al. 2005).
Black swallow-wort roots. Photo: Leslie J. Mehrhoff, University of Connecticut, Bugwood.org
Black swallow-wort seeds. Photo: Leslie J. Mehrhoff, University of Connecticut, Bugwood.org
Black swallow-wort seed pods. Photo: Leslie J. Mehrhoff, University of Connecticut, Bugwood.org
Pale swallow-wort seed pods and seeds. Photo: John M. Randall, The Nature Conservancy, Bugwood.org
Impacts
Black and pale swallow-wort are moderately aggressive. The extensive patches of swallow-wort may overgrow native plants and disrupt natural succession. Herbivores and grazers, such as deer, tend to avoid swallow-wort to feed on more palatable plants. Additionally, due to the tangling nature of swallow-wort, large patches can be difficult to walk through and may interfere with forest management and recreation.
Lower arthropod diversity and abundance have been observed on invasive pale swallow-wort when compared to native plants (Ernst and Cappuccino 2005). Bird populations and soil microbial communities may also be indirectly impacted when swallow-wort invades. Pale swallow-wort grows particularly well on limestone-based soils and is invading globally rare alvar habitats in New York (DiTommaso et al. 2005).
Studies have looked at the potential impact of swallow-wort on monarch butterflies (Danaus plexippus). Most monarch butterflies tend not to lay their eggs on the two swallow-wort species, but if they do the larvae cannot survive. Additionally, the butterflies may indirectly be threatened if swallow-wort outcompetes their preferred larval host, common milkweed (Asclepias syriaca) (DiTommaso and Losey 2003).
Identification
Pale and black swallow-wort look very similar when not in flower. Both species have long, oval, opposite leaves (about 3-4 inches long by 2-3 inches wide). The leaves of pale swallow-wort tend to be a slightly lighter shade of green than black swallow-wort. Flowers of both species are small and star-like. Pale swallow-wort has a pink to maroon flower with petals that have a narrow base and no hairs, whereas black swallow-wort’s are a deep purple-black and have petals with a wider base (about half as wide as they are long) and hairy inner petals. Both species produce seed pods that are 1.5″ to 3″ long with wind-distributed seeds inside. Swallow-worts possess a fibrous root system.
Flowers of black swallow-wort. Photo: Leslie J. Mehrhoff, University of Connecticut, Bugwood.org
Flowers of pale swallow-wort. Photo: Leslie J. Mehrhoff, University of Connecticut, Bugwood.org
Flowers and foliage of pale swallow-wort. Photo: Leslie J. Mehrhoff, University of Connecticut, Bugwood.orgLine drawing of black swallow-wort (Pale swallow-wort would look similar). Illustration: USDA PLANTS Database, USDA NRCS PLANTS Database, Bugwood.org
Prevention and Control
Mechanical and manual control:
Mechanical and manual control of swallow-wort are difficult due to the presence of a deep, fibrous root system. If you dig up either of these plants, try to get as much of the root crown as possible. For small stands, the plants and seed pods can be collected in plastic bags and destroyed or disposed of properly in a landfill before they release their seed. Collecting seed pods is not a long-term solution and may be impractical for large stands.
Mowing will not eradicate the plant, but can be utilized to prevent seed formation if timed correctly. If swallow-wort stands are cut too early, the plants can recover and still produce viable seed. It is suggested to mow when immature seed pods are just starting to develop and do not yet contain viable seeds (McKague and Cappuccino 2005). Mowing of swallow-wort after seed development is not recommended as it could aid in seed dispersal.
Early seed pods starting to develop on pale swallow-wort. Photo: Rob Routledge, Sault College, Bugwood.org
Chemical Control:
Chemical herbicides, such as glyphosate or triclopyr, may be used for control as foliar sprays or on cut stems (Weston et al. 2005). Recent work in central New York State has shown that applying these herbicides in late summer (i.e. mid- to late-August) to plants that have been mowed or trimmed in early July may provides the best results. The mowing of plants in early July also ensures that no viable seeds are produced prior to application of the herbicides (DiTommaso et al., unpublished). These chemicals work on living plants and need ample leaf surface area to absorb the herbicide to effectively kill the roots. For this reason, it is not recommended to apply them on seedlings soon after emergence. Results may been seen in one to two weeks, but repeated application may be needed. Check with your local extension office about herbicide use and permits needed for chemical application. As always, with any chemical, follow the instructions on the label.
Biological Control:
At this time, there are no approved biological control methods for swallow-wort. Biological controls for swallow-wort species are currently being investigated.
Occurrences
There have been reports of invasive black swallow-wort from Ontario, Canada south to Pennsylvania and from the northern Atlantic coast west to Missouri. Pale swallow-wort is most abundant in the lower Great Lakes Basin and has been observed in New York, Connecticut, Indiana, Massachusetts, Michigan, New Hampshire, New Jersey, and Pennsylvania as well as in parts of Canada.
New York Distribution Map
This map shows confirmed observations (green points) submitted to the NYS Invasive Species Database. Absence of data does not necessarily mean absence of the species at that site, but that it has not been reported there. For more information, please visit iMapInvasives.
A thicket of Japanese knotweed. Citation: Tom Heutte, USDA Forest Service, www.invasives.org
Background
Japanese knotweed (Polygonum cuspidatum Sieb. & Zucc.), a member of the buckwheat family, was introduced into the U.S. from Eastern Asia (Japan, China, Korea) as an ornamental on estates in the late-1800s. It has also been used as an erosion control plant. By the mid-1890s, it was reported near Philadelphia, PA, Schenectady, NY, and in New Jersey. Although once sold through seed and plant catalogs, by the late-1930s knotweed was already being viewed as a problematic pest. The plant, which can grow from three to 15 feet tall, has bamboo-like stems and is sometimes called Japanese bamboo. As with many invasive plants, knotweed thrives in disturbed areas and once established can spread rapidly, creating monoculture stands that threaten native plant communities. Japanese knotweed can tolerate deep shade, high temperatures, high soil salinity and drought. It is commonly found along streams and rivers, in low-lying areas, disturbed areas such as rights-of-way, and around old home and farmsteads.
Distribution
Japanese knotweed can be found in the U.S. in 42 states coast to coast except the arid Southwest, several of the deep South Gulf states and the highest of the Rocky Mountains. It is also in eight Canadian Provinces.
US Distribution of Japanese Knotweed. Map: USDA NRCS PLANTS Database
Identification
The plant is an upright, shrubby, herbaceous, woody-appearing perennial reaching heights of 10 to 15 feet (3 to 4.5 m). The stems are smooth, stout, and hollow. Where the leaves attach to the stem, the stem is swollen with a membranous sheath surrounding the joints. The leaves are broadly ovate (broad and rounded at the base and tapering toward the end), 3 to 6 inches (8 to 15 cm) long by 2 to 4 inches (5 to 10 cm) wide, alternating on stem, broadly oval to somewhat triangular or heart-shaped, pointed at the tip. The plant’s shoots come up from a network of spreading rhizomes. These horizontal roots can reach lengths of 65 feet (20 m) or more. Japanese knotweed has branched sprays of small greenish-white flowers from August to September. Although they have complementary male and female organs, those organs are vestigial and the flowers function unisexually. The plant develops small winged fruits Seeds: triangular, shiny, very small, about 1/10 inch (2.5 mm) long. Japanese knotweed spreads primarily by seed (transported by wind, water, animals, humans, or as a soil contaminant), stem fragments, and by shoots sprouting from its system of rhizomes.
Japanese knotweed showing oblate leaves and flowers. Jan Samanec, State Phytosanitary Administration, Bugwood.org
Jan Samanek, State Phytosanitary Administration, Bugwood.org – See more at: http://www.forestryimages.org/browse/detail.cfm?imgnum=5205100#sthash.WY0qNOzS.dpuf
Jan Samanek, State Phytosanitary Administration, Bugwood.org – See more at: http://www.forestryimages.org/browse/detail.cfm?imgnum=5205100#sthash.WY0qNOzS.dpuf
Jan Samanek, State Phytosanitary Administration, Bugwood.org – See more at: http://www.forestryimages.org/browse/detail.cfm?imgnum=5205100#sthash.WY0qNOzS.dpuf
Impacts
Japanese knotweed spreads rapidly, forming dense thickets that crowd and shade out native vegetation. This reduces species diversity, alters natural ecosystems, and negatively impacts wildlife habitat. The ground under knotweed thickets tends to have very little other growth. This bare soil is very susceptible to erosion, posing a particular threat to riparian areas. Once established, populations of Japanese knotweed are extremely persistent and hard to eradicate.
Prevention & Control
As with most invasive plants, Japanese knotweed can quickly inhabit disturbed ecosystems. One of the best ways to prevent its colonization is to ensure that disturbed habitats are rehabilitated with native vegetation before knotweed can invade.
Mechanical control
Single young plants can be pulled by hand depending on soil conditions and root development. If all of the root system isn’t removed, re-sprouting can occur. For small initial populations beyond single plants, or in environmentally sensitive areas where herbicides cannot be used, grubbing with a pulaski or similar tool to remove all of the roots after cutting back the standing vegetation can be an effective control measure. All parts of the removed plants should be bagged and disposed of in a secure location. It is best if knotweed not be mown or cut with weed trimmers as the pieces of the plant can easily get moved around and re-sprout, spreading, rather than controlling the plant.
Chemical control
Chemical controls for Japanese knotweed include application of glyphosate and triclopyr herbicides to freshly cut stems or to foliage. Before using chemicals to control any invasive plant, check with your local environmental or natural resources management agency or Cooperative Extension office to find out what chemicals are legal for use on knotweed in your area.
Catskill Regional Invasive Species Partnership (CRISP) Japanese knotweed video
New York Distribution Map
This map shows confirmed observations (green points) submitted to the NYS Invasive Species Database. Absence of data does not necessarily mean absence of the species at that site, but that it has not been reported there. For more information, please visit iMapInvasives.
Several species of honeysuckle found in NY are characterized as invasive, including: Morrow’s honeysuckle (Lonicera morrowii), Tatarian honeysuckle (Lonicera tatarica), Amur honeysuckle (Lonicera maackii), and Japanese honeysuckle (Lonicera japonica). To the non-botanist, native and invasive non-native honeysuckles appear very similar. One way to distinguish between native and invasive honeysuckles is by looking at the stems – native honeysuckles have solid stems while invasive honeysuckles have hollow stems.
Habitats
All four species are successful invaders of a similar range of habitats, including: abandoned fields; pastures; early successional, open canopy, and planted forests; along the edge of woodlots; floodplains; highway, railway and utility rights-of-way; open disturbed areas; vacant lots; edges of lawns; and, gardens. L. japonica can also be found in agricultural fields. All four grow best in full sun; L. japonica is the most shade-tolerant of the four, with L. tatarica and L. maackii being semi-shade tolerant.
Origin and Introduction
L. tatarica is native to Central Asia and Southern Russia and is believed to have been introduced into North America for ornamental purposes as early as the 1750s. L. japonica, – a native of China, Japan and Korea – was introduced for horticultural purposes in 1806 on Long Island; it was widely distributed as a garden plant through the early-1900s when it was finally recognized as a weed. L. maackii, also native to China, Japan and Korea, was introduced as seeds to arboreta throughout the U.S. in the late-1800s to determine whether the plant would grow in North America. This species of honeysuckle was utilized as a soil stabilization and wildlife planning until the mid-1980s and is still available for sale on-line. L. morrowii, a native of Japan, was imported to Massachusetts in the 1860s and was later released as an ornamental. All four species have escaped cultivation and are easily spread by birds.
Identification
L. morrowii, L. tatarica, and L. maackii), are perennial shrubs; L. japonica is a perennial woody vine (although its leaves can remain green throughout mild winters). The shrub forms range from 6 to 15 feet in height, while vines can reach 30 feet in length. The egg-shaped leaves range from 1 to 3 inches in length and are arranged oppositely along stems. Invasive honeysuckles begin flowering from May to June and bear small (less than 1 inch long), very fragrant tubular flowers ranging from creamy white through various shades of pink to crimson. L. morrowii and L. tatarica produce ¼ inch red berries from mid-summer through early-fall; L. maackii’s dark-red berries don’t ripen until late-fall; L. japonica produces dark-purple or black berries in the fall. Stems of all four are hollow.
Impacts
All three species can form very dense populations that can outcompete and suppress the growth of native plant species. These dense stands suppress the growth of other native species. L. maackii leafs out very early in spring, giving it a competitive advantage over native plants. L. japonica leaves are semi-evergreen allowing the plant to grow longer into the winter and giving it a competitive advantage over native vegetation. It shades out understory growth preventing the success of native understory plants and tree seedlings. Its vigorous vine growth covers native trees; the weight of the vine growth can bring down weak trees. By decreasing light availability to the understory, these invasive honeysuckles can alter habitats by depleting soil moisture and nutrients. The invasive honeysuckle berries do not contain the amount of fat and nutrients present in native honeysuckle berries; eating large amounts of the less nutritious invasive berries rather than native berries can have negative impacts on migrating.
Prevention and Control
Because these plants spread rapidly via birds eating seeds, control should be started in late-summer or early-fall before seeds are ready to be dispersed. In early stages of invasion, or in cases where populations are at low levels, hand removal of honeysuckle seedlings or young plants is a viable option when repeated annually. Systemic herbicides can be utilized in cases of heavy infestation. Specific state rules should be followed and the appropriate (low environment impact, legally labeled for control of these plants) herbicides should be used. For invasive honeysuckles growing in open habitats, prescribed burning may be an effective control alternative.
New York Distribution Map
This map shows confirmed observations (green points) submitted to the NYS Invasive Species Database. Absence of data does not necessarily mean absence of the species at that site, but that it has not been reported there. For more information, please visit iMapInvasives.











.jpg)













.jpg)